Peter F. Riederer und Rainer Burger, Würzburg
Wer Schokolade isst, weiß, was er sich Gutes tut. Schon der Anblick stimuliert die Sinne und dies wird durch Betasten, das Geräusch des Knackens, den Geruch und schließlich das Zergehen der Köstlichkeit auf der Zunge verstärkt. So ist es einfühlbar und nachvollziehbar, dass Schokolade die Psyche im Sinne eines Stimulus beeinflusst, euphorisierend wirkt und damit antidepressiven Charakter hat [28]. Ist es also nur Psychologie oder verbirgt sich in Schokolade ein pharmakologisches Prinzip, welches den Stimulus erklären lässt?
Ein Inhaltsstoff mit stimmungsaufhellenden Eigenschaften ist das Cannabinoid Anandamid, ein Abkömmling der Arachidonsäure. Um entsprechende Effekte auszulösen, müsste aber Schokolade in Kilogrammmengen konsumiert werden [38]. Ähnliches gilt für die Theobromin-Konzentration in Schokolade.
Häufig wird auch Serotonin als antidepressiv wirkender Stimulus genannt. Serotonin ist ein „biogenes Amin“, also eine Substanz, die biologisch aktiv ist. In der Tat ist die Serotoninhypothese der Depression [13], die im depressiven Pol ein Serotonindefizit an serotonergen Synapsen des Gehirns postuliert, Grundlage vieler antidepressiv wirkender Medikamente und speziell der selektiven Serotonin-Wiederaufnahmehemmer (SSRI).
Schokolade enthält in seinem Kakao-anteil eine Reihe biogener Amine, wie eben Serotonin, aber auch Dopamin, Tyramin, β-Phenylethylamin (PEA) und andere [29, 42]. Die Attraktivität, Serotonin aus diesem Pool herauszugreifen und als die Psyche stimulierende Substanz von Schokolade herauszuheben, beruht auf dem Gesagten.
Der von Wurtman und Wurtman [40] beschriebene Zusammenhang von Kohlenhydratmetabolismus, Blut-Tryptophankonzentration und Gehirn-Serotoninspiegel hat in der attraktiven Hypothese forcierten Kohlenhydratkonsums von Personen mit Serotonindefizit Widerhall gefunden und wird speziell bei „saisonaler Depression“ als basaler pathogener Mechanismus diskutiert. Parker et al. [28] widerlegen diese Hypothese aber sowohl aus psychiatrischer Sicht als auch basierend auf dem Gehalt an entsprechenden Inhaltsstoffen von Schokolade. Tabelle 1 gibt die wichtigsten Inhaltsstoffe von dunkler Schokolade an. Der Kohlenhydratanteil besteht vorwiegend aus dem Disaccharid Saccharose.
Tab. 1. Inhaltsstoffe in 100 g Schokolade mit 40% Kakaoanteil und der empfohlene Tagesbedarf1 [nach 22b, 37b]
|
Hauptinhaltsstoffe |
Menge [g] |
|
|
Kohlenhydrate |
47,0 |
|
|
Lipide |
30,0 |
|
|
Ballaststoffe |
15,0 |
|
|
Proteine |
5,3 |
|
|
Wasser |
0,9 |
|
|
Mineralstoffe und Spurenelemente (in Ionenform) |
Menge [mg] |
Empfohlene tägliche Zufuhrmenge [mg]3 |
|
Natrium |
19 |
550 |
|
Kalium |
397 |
2000 |
|
Magnesium |
100 |
300–400 |
|
Calcium |
63 |
1000 |
|
Eisen |
3,2 |
Männer: 10; Frauen4: 15 |
|
Kupfer |
1,4 |
1–1,5 |
|
Zink |
1,8 |
Männer: 10; Frauen: 7 |
|
Nickel |
0,374 |
k. A. |
|
Phosphor |
287 |
700 |
|
Chlorid |
100 |
830 |
|
Fluorid |
75 |
Männer: 3,8; Frauen: 3,1 |
|
Iodid |
5,5 |
0,2 |
1: nach Empfehlungen der Deutschen Gesellschaft für Ernährung; 2: Milchfreie Schokolade mit 40% Kakaomasse; 3: für Erwachsene; 4: Frauen im gebärfähigen Alter; k. A. keine Daten der DGE zur empfohlenen Zufuhrmenge vorhanden
Vergessen wird aber, dass Schokolade die Vorstufen dieser biogenen Amine in größerer Menge enthält. Das sind Aminosäuren wie die aromatischen Aminosäuren Phenylalanin, Tyrosin, Dihydroxyphenylalanin (DOPA), Tryptophan und 5-Hydroxytryptophan (Tab. 2 und Abb. 1). Der Zusammenhang sollte bei der Überlegung eines stimulierenden Effekts biogener Amine nicht übersehen werden, da aus den Aminosäurevorstufen nach Aufnahme von Schokolade biogene Amine durch die aromatische Aminosäuredecarboxylase gebildet werden können.
Tab. 2. Aminosäuren und deren biogene Amine
|
Aminosäuren |
Biogenes Amin |
|
Tryptophan |
Tryptamin, Kynurenin |
|
5-Hydroxytryptophan |
Serotonin, Melatonin |
|
Phenylalanin |
b-Phenylethylamin |
|
Tyrosin |
Tyramin |
|
L-DOPA |
Dopamin, Noradrenalin |
|
Histidin |
Histamin |
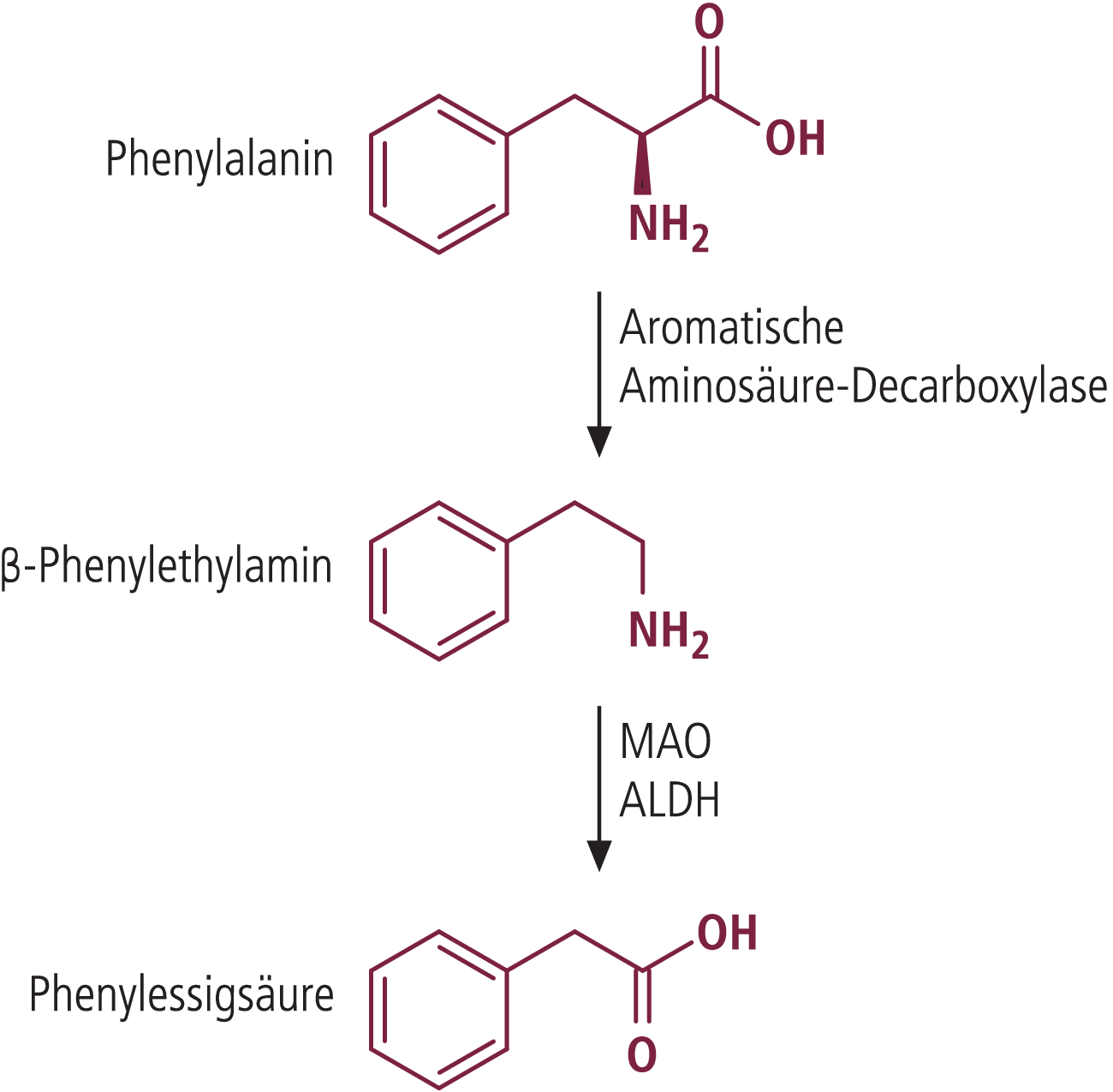
Abb. 1. Biosynthese und Abbau von b-Phenylethylamin. MAO: Monoaminoxidase; ALDH: Aldehyddehydrogenase
Da es an entsprechenden aktuellen Daten mangelt, haben wir in einem ersten Schritt den Aminosäuregehalt und die Konzentration biogener Amine in kommerziell erhältlicher Schokolade untersucht.
Methoden
Kommerziell erhältliche Schokolade mit unterschiedlichem Anteil an Kakaobestandteilen (37, 40, 50, 72, 75 und 85%) wurde per HPLC auf ihren Gehalt an 5-Hydroxytryptophan, Serotonin (5-Hydroxytryptamin, 5-HT), 5-Hydroxyindolessigsäure (5-HIES), 3,4-Dihydroxyphenylalanin (DOPA), Dopamin (DA), 3,4-Dihydroxyphenylessigsäure (DOPAC) und Homovanillinsäure (HVA) sowie verschiedener Aminosäuren untersucht. Die ausführliche Methodenbeschreibung ist auf der Website der „Psychopharmakotherapie“ beim Inhaltsverzeichnis dieses Hefts dargestellt: www.ppt-online.de > Inhalt > Jahrgang 2009 > Heft 1
Resultate
100 g kommerziell erhältliche Schokolade enthält, abhängig vom Gehalt an Kakaotrockenmasse, unterschiedliche Konzentrationen der einzelnen Aminosäuren, speziell aber von neutralen Aminosäuren (aromatische und verzweigtkettige Aminosäuren Valin, Leucin und Isoleucin), biogenen Aminen und deren Metaboliten. Schokolade mit einem hohen Anteil an Kakaotrockenmasse zeigt wesentlich höhere Konzentrationen der meisten dieser Bestandteile als Schokolade mit einem niedrigeren Anteil an Kakaotrockenmasse (Tab. 3).
Tab. 3. Anteile [µmol/g Schokolade] von Aminosäuren, biogenen Aminen und Metaboliten in Schokolade (Beispiele)
|
Aminosäure |
Kakaotrockenmasse [%]* |
|||||
|
37 |
40 |
50 |
72 |
75 |
85 |
|
|
Phenylalanin# |
15,08 |
14,57 |
14,31 |
23,38 |
21,26 |
20,11 |
|
PEA (b-Phenylethylamin) |
17,03 |
18,65 |
16,05 |
28,61 |
25,11 |
24,12 |
|
Tyrosin# |
17,17 |
15,63 |
15,11 |
19,34 |
26,53 |
26,72 |
|
DOPA (3,4-Dihydroxyphenylalanin)# |
1,01 |
1,18 |
1,09 |
1,25 |
1,40 |
1,46 |
|
Dopamin |
12,11 |
13,71 |
20,47 |
48,21 |
40,17 |
43,79 |
|
DOPAC (3,4-Dihydroxyphenylessigsäure) |
9,52 |
12,50 |
21,18 |
32,68 |
41,67 |
48,64 |
|
HVA (Homovanillinsäure) |
19,52 |
34,01 |
55,74 |
45,81 |
50,32 |
52,95 |
|
Tryptophan# |
26,67 |
31,98 |
26,59 |
46,34 |
45,01 |
38,07 |
|
5-OH-Tryptophan# |
2,65 |
2,66 |
3,43 |
3,83 |
4,41 |
3,95 |
|
5-HT (Serotonin) |
5,87 |
12,40 |
16,05 |
20,17 |
66,02 |
60,41 |
|
5-HIES (5-Hydroxyindolessigsäure) |
4,75 |
6,07 |
8,04 |
12,65 |
13,18 |
12,78 |
|
Valin## |
20,97 |
21,34 |
21,32 |
24,99 |
22,38 |
25,06 |
|
Leucin## |
20,46 |
19,25 |
19,68 |
30,75 |
29,95 |
28,22 |
|
Isoleucin## |
21,63 |
19,23 |
19,36 |
40,85 |
39,79 |
35,85 |
|
Asparagin |
20,34 |
18,53 |
19,30 |
32,41 |
28,26 |
25,96 |
|
Aspartat |
15,69 |
17,46 |
15,14 |
21,22 |
22,69 |
23,49 |
|
Glutamin |
18,78 |
18,77 |
18,50 |
19,02 |
19,33 |
18,61 |
|
Glutamat |
27,67 |
34,03 |
30,02 |
44,84 |
34,10 |
29,12 |
|
Serin |
52,26 |
42,82 |
43,87 |
51,22 |
44,93 |
44,95 |
|
Histidin |
11,17 |
13,69 |
12,68 |
13,33 |
13,35 |
13,26 |
|
Glycin |
27,98 |
30,61 |
27,54 |
45,39 |
41,70 |
37,67 |
|
Threonin |
18,65 |
22,38 |
19,58 |
27,19 |
31,21 |
28,69 |
|
Arginin |
12,58 |
9,71 |
8,24 |
18,34 |
18,23 |
16,90 |
|
Alanin |
44,25 |
43,21 |
39,49 |
59,00 |
59,41 |
58,72 |
|
Methionin |
38,58 |
38,68 |
38,43 |
60,23 |
55,61 |
52,24 |
|
Lysin |
20,78 |
17,94 |
18,69 |
42,20 |
44,28 |
36,39 |
*Kommerziell erhältliche Schokolade einer Produktserie
37: ≤37% Kakao, ≤ 20% Milchanteil # aromatische Aminosäuren
40: ≤40% Kakao, 23% Milchanteil ##verzweigtkettige Aminosäuren
50: ≤50% Kakao, ≤18% Milchanteil Summe aus aromatischen und verzweigtkettigen
72: ≤72% Kakao, kein Milchanteil Aminosäuren: neutrale Aminosäuren
75: ≤75% Kakao, kein Milchanteil
85: ≤85% Kakao, kein Milchanteil
Diskussion
Folgende Fragen sind also zu klären, um einen möglichen stimulierenden, das heißt einen sich im Gehirn abspielenden Vorgang in Nervenzellen verständlich zu machen:
Was passiert nach der Einnahme von Schokolade in der Peripherie, also außerhalb des zentralen Nervensystems?
Die Rolle der „neutralen“ Aminosäuren. Der Aminosäurepool des Plasmas sowie aller Organe wird durch Aufnahme und Abgabe von Aminosäuren aus dem peripheren Stoffwechsel der Organe, durch Ein- und Abbau von Eiweiß und durch die Nahrung gesteuert. Die Konzentration der aromatischen und verzweigtkettigen Aminosäuren liegt in der Größenordnung von 10 bis 340 µmol/l Plasma. Tryptophan ist die einzige Aminosäure, die zu etwa 90% an Albumin gebunden ist. Bemerkenswert ist auch, dass Tryptophan zum allergrößten Teil im sogenannten „Kynurenin- Stoffwechsel“ metabolisiert wird. Im Gehirn wird Tryptophan unter anderem in serotoninerge Nervenzellen aufgenommen. Da die Tryptophanhydroxylase 2 (nur diese Isoform ist für die Serotoninsynthese im Gehirn verantwortlich [22]) aber immer mit dem Substrat gesättigt ist, spielt eine etwaige Konzentrationserhöhung von Tryptophan in serotoninergen Neuronen für die Serotoninsynthese keine Rolle.
Die Rolle der biogenen Amine. Serotonin kann auch außerhalb des zentralen Nervensystems aus Tryptophan gebildet werden. Der Großteil wird in den Blutplättchen (Thrombozyten) gespeichert. Freies Plasma-Serotonin kann die Blut-Hirn-Schranke nicht passieren, da Monoaminoxidase (MAO) A der Epithelzellen Serotonin sofort zu 5-Hydroxyindolessigsäure abbaut. Eine erhöhte Serotonin-Plasmakonzentration hat daher keine Bedeutung für die Gehirnfunktion.
Auch die biogenen Amine Tyramin, Dopamin, Tryptamin können in der Peripherie aus den entsprechenden Aminosäuren biosynthetisiert werden. Sie sind aber ebenfalls bevorzugte Substrate der MAO-A und werden daher, wie auch Serotonin, an der Blut-Hirn-Schranke durch MAO-A abgebaut.
β-Phenylethylamin wird peripher aus Phenylalanin biosynthetisiert. Freies Plasma-Phenylethylamin kann die Blut-Hirn-Schranke gut passieren, da es das bevorzugte Substrat der MAO-B ist, die in den Endothelzellen der Kapillaren und daher in der Blut-Hirn-Schranke nicht vorkommt. Es wird auch im Gehirn aus der Aminosäure Phenylalanin gebildet. Zusätzlich kann es, wie bei Schokolade auch, aus der Nahrung direkt aufgenommen werden. Veränderungen der Plasmakonzentration von β-Phenylethylamin haben daher direkte Auswirkungen auf neuronale Funktionen des Gehirns.
Es ist daher sehr wahrscheinlich, dass nur β-Phenylethylamin bei verstärktem Angebot, also beispielsweise nach Konsum von Schokolade mit hohem Kakaoanteil, vermehrt in das Gehirn aufgenommen wird und dort neuronale Aktivität entfaltet.
Wie werden biogene Amine und Aminosäuren in das Gehirn aufgenommen?
Nimmt man also 100 g Schokolade zu sich und damit neben den Aminosäuren aus dem Kakaoanteil eventuell auch Milcheiweiß und Aminosäuren der Milch, kann das den Aminosäurepool des Plasmas selbst, nach Berücksichtigung der Verluste durch Resorption und Metabolismus von Aminosäuren im Magen-Darm-Trakt, durch eine Konzentrationserhöhung beeinflussen. Der freie, nicht an Albumin gebundene Anteil von Tryptophan folgt dieser Gegebenheit, da zwischen freiem und gebundenem Anteil ein Gleichgewicht besteht [2, 15, 26]. Aromatische und verzweigtkettige Aminosäuren (Valin, Leucin und Isoleucin), zusammen auch „neutrale Aminosäuren“ genannt, werden über einen gemeinsamen aktiven Transportmechanismus in das Gehirn aufgenommen [33]. Dieser aktive Transport unterliegt einem „Wettbewerb“. Aminosäuren, die in höherer Konzentration angeboten werden, werden verstärkt aufgenommen und behindern daher die Aufnahme geringer konzentrierter anderer neutraler Aminosäuren (z.B. bei der Levodopa-Therapie der Parkinson-Krankheit).
Günstige vaskuläre Eigenschaften von Kakao, bedingt durch Flavanole, unterstützen die neuronalen Wirkungen von Schokolade [18, 19].
Tabelle 3 zeigt deutlich, dass das Verhältnis der Aminosäurekonzentration von Schokolade unabhängig von der Konzentration der Kakaobestandteile ist und in etwa gleich bleibt. Es ist daher nicht anzunehmen, dass sich die Aufnahmerate der neutralen Aminosäuren in das Gehirn beim Konsum von Schokolade mit unterschiedlichem Kakaogehalt entscheidend verändert.
Wie wirken biogene Amine nach Aufnahme in das Gehirn an entsprechenden Synapsen?
Die Kernfrage ist: Kann b-Phenylethylamin im Gehirn eine „stimulierende Wirkung“ entfalten?
Einmal im Gehirn präsent, durchdringt β-Phenylethylamin wegen seiner guten lipophilen Eigenschaften leicht Membranen, auch diejenigen von Nervenzellen. Die Strukturverwandtschaft zu den Molekülen Dopamin und Amphetamin lässt ahnen, dass β-Phenylethylamin physiologische Wirkung generieren sollte. In der Tat sind für β-Phenylethylamin Dopamin- und Noradrenalin-freisetzende Eigenschaften beschrieben worden [20] und es scheint möglicherweise direkte agonistische Effekte an Dopamin- und Serotoninrezeptoren zu haben [1, 23, 36]. Experimentelle Studien weisen darauf hin, dass β-Phenylethylamin Stereotypien auslöst [31, 34], die durch MAO-B-Hemmer verstärkt [24] und durch Neuroleptika antagonisiert [10, 25] werden. Die Konzentration von β-Phenylethylamin im Gehirn wird durch den MAO-B-Hemmer Selegilin (L-Deprenyl) erhöht, wie Post-mortem-Untersuchungen von Patienten ergeben haben [32]. Neuere tierexperimentelle Untersuchungen mit Affen zeigen, dass MAO-B-Hemmer wie Selegilin psychomotorisch stimulierende Effekte von β-Phenylethylamin potenzieren [5, 35].
Neuronal modulierende postsynaptische Eigenschaften wurden von Reynolds [19] diskutiert. In der Tat zeigen neuere Untersuchungen, dass es für biogene Amine wie β-Phenylethylamin sogenannte „Trace Amine Associated Receptors“ (TAAR) gibt [11, 12], die auf direkte Stimulation einer Rezeptorkaskade durch „Spurenamine“ hinweisen (Tab. 4). Eine TAAR-1-Aktivierung durch β-Phenylethylamin hemmt die Wiederaufnahme und den induzierten Efflux von [3H]-Dopamin, [3H]-Noradrenalin und [3H]-Serotonin in transfizierten Zellen. In striatalen Synaptosomen des Affengehirns hemmt β-Phenylethylamin signifikant, die Wiederaufnahme und den induzierten Efflux von [3H]-Dopamin und [3H]-Serotonin [41].
Tab. 4. Potenzielle neurochemische und pharmakologische Eigenschaften von b-Phenylethylamin
|
Blut-Hirn-Schrankengängigkeit |
|
Lipophiles Amin |
|
Strukturelle Ähnlichkeit zu Dopamin und Amphetamin |
|
Wiederaufnahmehemmer von Dopamin, Noradrenalin und Serotonin |
|
Verstärkung der präsynaptischen Freisetzung von Dopamin, Noradrenalin und Serotonin |
|
Stimulation von Dopamin-Autorezeptoren |
|
Dopamin-/Serotonin-Antagonist? |
|
Unterdrückung des inhibitorischen postsynaptischen Potenzials |
|
Stimulierung von TAA-Rezeptoren |
|
Auslöser von Sterotypien in Tierexperimenten |
|
Verstärkung der Effekte durch MAO-Inhibitoren |
|
Antagonismus der Verhaltensänderungen durch Neuroleptika |
|
Stimulation der Psychomotorik |
|
Antidepressive Wirkungen |
Obwohl Spurenamine wie β-Phenylethylamin dopaminerge Autorezeptoren durch gesteigerten Efflux von neu synthetisiertem Dopamin indirekt aktivieren [21], weisen die Untersuchungen von Xie und Miller [39] darauf hin, dass β-Phenylethylamin die Monoamintransporterfunktion durch Interaktion mit TAAR-1, nicht aber durch eine solche über Autorezeptoren, beeinflusst. Der Dopamintransporter ist für die striatale Dopamin-freisetzende und hyperlokomotorische Wirkung von β-Phenylethylamin notwendig. Dagegen scheint die hemmende Wirkung auf die β-Phenylethylamin-induzierte Hyperaktivität und Stereotypien in Dopamintransporter-Knock-out-Mäusen durch andere Mechanismen verursacht zu werden [37].
Spurenamine wie β-Phenylethylamin reduzieren die GABA-B-Antwort in dopaminergen Nervenzellen. Diese Unterdrückung des inhibitorischen synaptischen Potenzials in Neuronen des zentralen Nervensystems durch β-Phenylethylamin unterstreicht dessen wichtige Rolle als Neuromodulator [14] (Tab. 4).
Es ist daher nicht verwunderlich, dass β-Phenylethylamin mit psychischen Erkrankungen im Allgemeinen [32] und Schizophrenie im Besonderen [4, 16, 30] in Verbindung gebracht wurde. Aber auch von Schokoladesucht als Persönlichkeitsmarker Depressiver wurde berichtet [27].
Ein gemütstimulierender Effekt von L-Phenylalanin bei Parkinson-Patienten mit Depressionen wurde von Birkmayer [6] beschrieben. Antidepressive Effekte bei Patienten mit Depressionen wurden von Birkmayer et al. [7], Fischer et al. [17] und Beckmann et al. [3] (D,L-Phenylalanin) beschrieben.
Für die Kombination aus L-Phenylalanin (250 mg/Tag) und dem MAO-B-Hemmer Selegilin (5–10 mg/Tag) haben Birkmayer et al. [8, 9] in einer offenen Studie sehr gute antidepressive klinische Effekte nachgewiesen, die die Autoren auf einen pharmakologischen Mechanismus von β-Phenylethylamin bezogen.
Antriebssteigernde und gemütsverbessernde Effekte von β-Phenylethylamin haben Pharmakologen dazu verleitet, β-Phenylethylamin als „endogenes Amphetamin“ zu bezeichnen, obwohl die pharmakologischen Eigenschaften beider Substanzen durchaus unterscheidbar sind.
Die Wirkung von β-Phenylethylamin scheint von der Gegenwart der Catecholamine, speziell Dopamin, abzuhängen [siehe aber 11, 12]. Darauf können auch die guten klinischen Effekte des MAO-B-Hemmers Selegilin als Anti-Parkinsonmittel bezogen werden, da der Abbau von β-Phenylethylamin durch Selegilin komplett gehemmt wird und β-Phenylethylamin endogenes und durch Levodopa substituiertes Dopamin verstärkt in dopaminerge Synapsen freisetzen kann.
Bemerkenswert ist daher, dass Selegilin zu einer Steigerung der β-Phenylethylamin-Konzentration im Gehirn von Parkinsonpatienten führt, die auch post mortem noch nachweisbar ist [32]. Im Rahmen einer Nachfolgestudie sollen daher Plasmakonzentrationen von β-Phenylethylamin nach Genuss von Schokolade gemessen werden. In diesem Zusammenhang sind Beobachtungen von Wolz et al. [39] von Interesse, die zeigen, dass der Schokoladekonsum von Parkinsonkranken erhöht ist. Derzeit ist eine klinische Studie mit dem Ziel, die Plasmakonzentration von β-Phenylethylamin bei Parkinsonkranken nach Einnahme einer bestimmten Menge 70%iger Schokolade zu bestimmen, in Arbeit (Reichmann et al., persönliche Mitteilung).
Alle diese Untersuchungen weisen darauf hin, dass β-Phenylethylamin ein nicht zu unterschätzendes biogenes Amin ist, das neuropsychiatrische Mechanismen beeinflusst oder selbst direkt ausübt. Diese werden auf der Verhaltensebene durch eine Stimulation motorischen Verhaltens sowie in „euphorisierender“ und stimmungsaufhellender Wirkung repräsentiert. Eventuell ist es also so, dass diese multiplen biochemischen und pharmakologischen Eigenschaften von β-Phenylethylamin durch psychologische Effekte der Vermarktung und durch die eingangs beschriebene Biopsychologie von der Art der Schokolade, Ansehen der Schokolade, Riechen, Hören und Tasten synergistisch beeinflusst werden. Somit erfüllt Schokolade viele Kriterien eines Pharmakons, basierend auf Inhaltsstoffen der Kakaobohnen, die je nach Herkunft unterschiedlich zusammengesetzt sein können.
Keiner der psychoaktiven Inhaltsstoffe von Schokolade wie Anandamid, Theobromin, Coffein, Tyramin, Tryptophan und b-Phenylethylamin erreicht eine Konzentration, die ausreichen würde, um Schokolade als „Stimulans“ zu definierten. Schokolade vermittelt hedonistische Belohnung durch die Befriedigung von „Verlangen“ [28] und b-Phenylethylamin mag dafür der geeignete „Stimulus“ sein.
Literatur
1. Antelman SM, Edwards DJ, Lin M. Phenylethylamine: evidence for a direct, postsynaptic dopamine-receptor stimulating action. Brain Res 1977;127:317–22.
2. Axelrod J, Saavedra J. Octopamine, phenylethanolamine, phenylethylamine and tryptamine in the brain. In: Ciba Foundation Symposium 22 (new series) (eds.). Aromatic Amino Acids in the Brain. Amsterdam, London, New York: Elsevier, 1974:51–66.
3. Beckmann H, Strauss MA, Ludolph E. DL-Phenylalanine in depressed patients: an open study. J Neural Transm 1977;41:123–34.
4. Beckmann H, Reynolds GP, Sandler M, Waldmeier P, et al. Phenylethylamine and phenylacetic acid in CSF of schizophrenics and healthy controls. Arch Psychiatr Nervenkr 1982;232:463–71.
5. Bergman J, Yasar S, Winger G. Psychomotor stimulant effects of beta-phenylethylamine in monkeys treated with MAO-B inhibitors. Psychopharmacology (Berl) 2001;159:21–30.
6. Birkmayer W. Experimentelle Befunde und neue Aspekte bei extrapyramidalen Erkrankungen. Z Nervenheilkunde (Wien) 1966;13:128–39.
7. Birkmayer W, Neumayer E, Stöckl W, Weiler G. Defizit essentieller Aminosäuren bei endogener Depression. Wien Klin Wochenschr 1968;80:832–4.
8. Birkmayer W, Riederer P. Die Parkinson-Krankheit. Biochemie, Klinik, Therapie. Wien, New York: Springer Verlag, 1980.
9. Birkmayer W, Riederer P. Linauer W, Knoll J. L-Deprenyl plus L-Phenylalanine in the treatment of depression. J Neural Transm 1984;59:81–7.
10. Borison RI, Hafdala HS, Diamond BI. Chronic phenylethylamine stereotypy in rats: a new animal model for schizophrenia. Life Sci 1977;21:117–22.
11. Borowsky B, Adham N, Jones KA, Raddatz R, et al. Trace amines: identification of a family of mammalian G protein-coupled receptors. Proc Natl Acad Sci U S A 2001;98: 8966–71.
12. Bunzow JR, Sonders MS, Arttamangkul S, Harrison LM, et al. Amphetamine, 3,4-methylenedioxymethamphetamine, lysergic acid diethylamide, and metabolites of the catecholamine neurotransmitters are agonists of a rat trace amine receptor. Mol Pharmacol 2001;60:1181–8.
13. Coppen A. The biochemistry of affective disorders. Br J Psychiatry 1967;113:1237.
14. Federici M, Geracitano R, Tozzi A, Longone P, et al. Trace amines depress GABA B response in dopaminergic neurons by inhibiting G-betagamma-gated inwardly rectifying potassium channels. Mol Pharmacol 2005;67: 1283–90.
15. Fernstrom JD, Madras BK, Munro HN, Wurtman RJ. Nutritional control of the synthesis of 5-hydroxytryptamine in the brain. In: Ciba Foundation Symposium 22 (new series) (eds.). Aromatic Amino Acids in the Brain. Amsterdam, London, New York: Elsevier, 1974:153–73.
16. Fischer E, Spatz H, Heller B, Reggiani H. Phenylethylamine content of human urine and rat brain, its alteration in pathological conditions and after drug administration. Experientia 1972;28:307.
17. Fischer E, Heller B, Nachon M, Spatz H. Therapy of depression by phenylalanine. Arzneimittelforschung (Drug Res) 1975;25:132.
18. Fisher ND, Hollenberg NK. Aging and vascular responses to flavanol-rich cocoa. J Hypertens 2006;24:1575–80.
19. Fisher ND, Sorond FA, Hollenberg NK. Cocoa flavanols and brain perfusion. J Cardiovasc Pharmacol 2006;47(Suppl 2):S210–4.
20. Fuxe K, Grobecker H, Jonsson G. The effect of β-phenethylamine on central and peripheral monoamine containing neurons. Eur J Pharmacol 1967;2:200–7.
21. Geracitano R, Federici M, Prisco S, Bernardi G, et al. Inhibitory effects of trace amines on rat midbrain dopaminergic neurons. Neuropharmacology 2004;46:807–14.
22. Gutknecht L, Waider J, Kraft S, Kriegebaum C, et al. Deficiency of brain 5-HT synthesis but serotonergic neuron formation in Tph2 knock-out mice. J Neural Transm 2008;115:1127–32.
22b. Hahn A, Ströhle A, Wolters M. Ernährung. 2. Aufl. Stuttgart: Wiss. Verl.-Ges., 2006.
23. Jackson DM. Beta-phenylethylamine and locomotor activity in mice. Interaction with catecholaminergic neurones and receptors. Arzneimittelforschung 1975;25:622–6.
24. Moja EA, Stoff DM, Gillin JC, Wyatt RJ. Dose-response effects of β-phenethylamine on stereotyped behaviour in pargyline-pretreated rats. Biol Psychiatr 1976;11:731–42.
25. Moja EA, Stoff DM, Gillin JC, Wyatt FJ. Neuroleptics attenuate stereotyped behaviour induced by β-phenethylamine in rats. Biol Psychiatr 1978;13:291–5.
26. Munro HN. Control of plasma amino acid concentrations. In: Ciba Foundation Symposium 22 (new series) (eds.). Aromatic Amino Acids in the Brain. Amsterdam, London, New York: Elsevier, 1974:5–24.
27. Parker G, Crawford J. Chocolate craving when depressed: a personality marker. Br J Psychiatry 2007;191:351–2.
28. Parker G, Parker I, Brotchie H. Mood state effects of chocolate. J Affect Disord 2006;92:149–59.
29. Pastore P, Favaro G, Badocco D, Tapparo A, et al. Determination of biogenic amines in chocolate by ion chromatographic separation and pulsed integrated amperometric detection with implemented wave-form at Au disposable electrode. J Chromatogr A 2005;1098: 111–5.
30. Potkin SG, Karoum F, Chuang LW, Cannon-Spoor HE, et al. Phenylethylamine in paranoid chronic schizophrenia. Science 1979;206:159.
31. Randrup A, Munkvad I. Dopa and other naturally occurring substances as causes of stereotypy and rage in rats. Acta Psychiatr Scand 1966;42(Suppl 191):193–9.
32. Reynolds GP, Riederer P, Sandler M. 2-Phenylethylamine and amphetamine in human brain: effects of L-deprenyl in Parkinson’s disease. Br Biochem Soc Trans 1978;7:143–5.
33. Riederer P. L-dopa competes with tyrosine and tryptophan for human brain uptake. Nutr Metab 1980;24:417–23.
34. Sabelli HC, Vazquez AJ, Flavin D. Behavioural and electrophysiological effects of phenylethanolamine and 2-phenylethylamine. Psychopharmacologia 1975;42:117–25.
35. Shimazu S, Miklya I. Pharmacological studies with endogenous enhancer substances: beta-phenylethylamine, tryptamine, and their synthetic derivatives. Prog Neuropsychopharmacol Biol Psychiatry 2004;28:421–7.
36. Sloviter RS, Connor JD, Drust EG. Serotonergic properties of β-phenethylamine in rats. Neuropharmacology 1980;19:1071–4.
37. Sotnikova TD, Budygin EA, Jones SR, Dykstra LA, et al. Dopamine transporter-dependent and -independent actions of trace amine beta-phenylethylamine. J Neurochem 2004;91:362–73.
37b. Souci SW, Fachmann W, Kraut H. Die Zusammensetzung der Lebensmittel, Nährwert-Tabellen. 7. Aufl. Stuttgart: Wiss. Verl.-Ges., 2008.
38. Tomaso di E, Beltramo M, Piomelli D. Brain cannabinoids in chocolate. Nature 1996;382:677–8.
39. Wolz M, Kaminski A, Löhle M, Koch R, et al. Chocolate consumption is increased in Parkinson’s disease: Results from a self-questionnaire study. (submitted).
40. Wurtman RJ, Wurtman JJ. Carbohydrates and depression. Sci Am 1989;260:68–75.
41. Xie Z, Miller GM. Beta-phenylethylamine alters monoamine transporter function via trace amine-associated receptor 1: implication for modulatory roles of trace amines in brain. J Pharmacol Exp Ther 2008;325:617–28.
42. Ziegleder G, Stojacic E, Stumpf B. Occurrence of beta-phenylethylamine and its derivatives in cocoa and cocoa products. Z Lebensm Unters Forsch 1992;195:235–8.
Prof. Dipl.-Ing. Dr. Peter F. Riederer, Rainer Burger, Klinische Neurochemie, Klinik für Psychiatrie, Psychosomatik und Psychotherapie, Füchsleinstr. 15, 97080 Würzburg, E-Mail: peter.riederer@mail.uni-wuerzburg.de
Is chocolate a psychotropic drug? – the role of b-phenylethalymine as psychostimulus
Chocolate contains a variety of substances, which by theoretical means are able to influence mood. However, based on the discussion in this article only β-phenylethylamine, a biogenic amine which crosses the blood-brain barrier very easily and prossesses properties similar but not identical to amphetamine, is able to influence transmitter metabolism and function. As this seems to be possible after consumption of about 100 g (black) chocolate, an influence on psyche, mood and activity seems to reflect such transmitter changes which are related to β-phenylethylamine. While it cannot be ruled out that psychological effects, like smelling, tasting or hearing/braking chocolate are of importance in its mood enhancing properties β-phenylethylamine may be a synergistically influencing factor.
Psychopharmakotherapie 2009; 16(01)