Walter E. Müller, Worms
Kognitive Dysfunktion und Depression
Sonderstellung innerhalb des depressiven Syndroms
Die unipolare Depression mit einer weltweiten Lebenszeit-Prävalenz von über 10% gehört schon seit vielen Jahren in der Liste der Erkrankungen, die zu schweren Beeinträchtigungen des Patienten führen (burden of disease), zu den Störungsbildern mit dem höchsten Beeinträchtigungsgrad. Zur Behandlung dieser Erkrankung, die oft phasenweise, aber auch häufig chronifizierend mit zunehmender Anzahl von depressiven Episoden verläuft, steht eine Reihe von medikamentösen und verhaltenstherapeutischen Therapien zur Verfügung, die aber nicht jedem Patienten primär helfen und die die Suche nach neueren und besseren therapeutischen Interventionen immer noch nötig erscheinen lassen [50, 62]. Im Zentrum des klinischen Bildes der Erkrankung steht das depressive Syndrom, dessen extreme Varianz mit psychischen, psychomotorischen und somatischen Symptomen auch im Zeitalter eher reduktionistischer standardisierter Diagnosen im Rahmen der ICD-10 oder der DSM-IV-R bzw. DSM-5 immer noch eine diagnostische Herausforderung darstellt. Wie von Laux und Goemann [50] ausgeführt, ist es immer noch irritierend, dass depressive Patienten mit identischer Diagnose eine extrem unterschiedliche klinische Symptomatik zeigen können. Dies hat wiederholt die Frage aufgeworfen, inwieweit die Erkrankung tatsächlich eine Entität darstellt oder ob das, was heute als depressive Störung diagnostiziert wird, sich nicht doch letztlich aus verschiedenen Grunderkrankungen zusammensetzt. Diese sehr komplexe Problematik weiter zu thematisieren, würde den Rahmen der vorliegenden Arbeit weit sprengen. Ein wichtiges Argument für die Hypothese eines sehr breiten depressiven Syndroms im Rahmen nur einer depressiven Grunderkrankung ist das mehr oder weniger zeitlich gemeinsame Auftreten der breiten Symptomatik, sowohl zu Beginn einer depressiven Episode wie auch nach deren Abklingen, entweder durch den natürlichen Verlauf oder durch eine therapeutische Intervention. Auch Befunde zur medikamentösen Therapie unterstützen die Hypothese eines breiten depressiven Syndroms: Heute gibt es eine ganze Reihe von sehr selektiven Antidepressiva mit spezifischen Angriffspunkten in zum Teil nur einem der drei aminergen Transmittersysteme (Abb. 1), die wir mit der Depression in Verbindung bringen. Trotz dieser sehr unterschiedlichen pharmakologischen Interventionsmöglichkeiten ist es bis jetzt nicht überzeugend gelungen, das therapeutische Ansprechen auf die individuellen Antidepressiva bestimmten Symptomenkomplexen innerhalb des depressiven Syndroms mit prädiktiver Sicherheit zuzuordnen [21]. Zwar findet man in den meisten Übersichten ähnliche Grafiken wie in Abbildung 1, die suggerieren, dass bestimmte Aspekte des depressiven Syndroms eher bestimmten Neurotransmittersystemen unseres Gehirns zuzuordnen sind und damit eher selektiv auf für diese Systeme spezifische Antidepressiva ansprechen sollten [94]. Bei kritischer Betrachtung muss man aber akzeptieren, dass diese Schemata eher didaktischen Wert als praktische Umsetzbarkeit aufweisen. Die meisten Leitlinien gehen davon aus, dass alle Antidepressiva mehr oder weniger symptomunabhängig das depressive Syndrom in seiner ganzen Ausprägung verbessern. Dies schlägt sich auch darin nieder, dass man heute fordert, dass ein klinisch akzeptables Antidepressivum diese symptomatische Breite im Rahmen depressiver Erkrankungen zeigen muss.
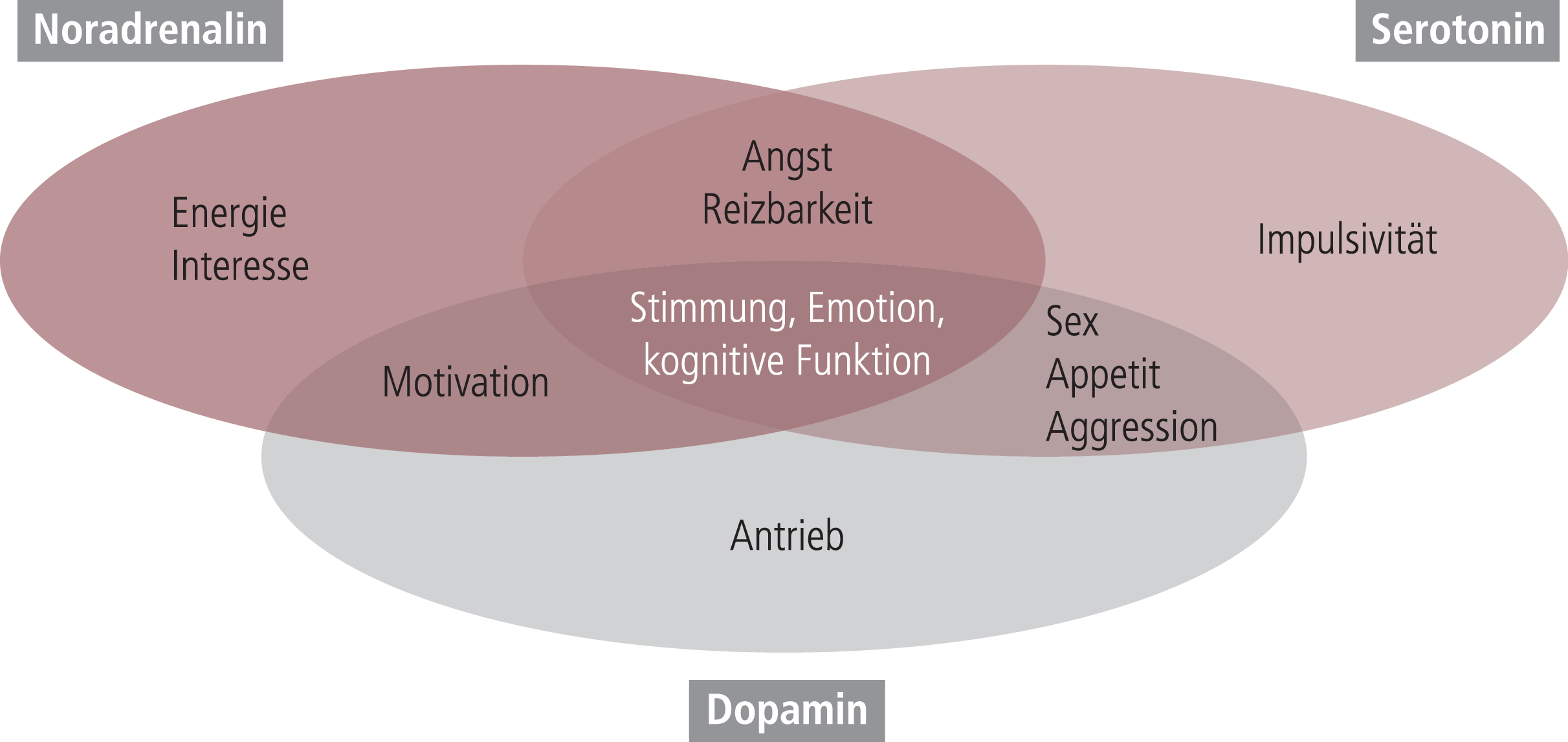
Abb. 1. Schematische Darstellung der möglichen funktionellen Bedeutung der drei zentralen aminergen Transmittersysteme für die einzelnen Symptombereiche des depressiven Syndroms. Bemerkenswert ist die große Überlappung besonders auch im Bereich der kognitiven Symptome.
Die großen Schwierigkeiten, mit den heutigen Antidepressiva bestimmte Symptome des depressiven Syndroms gezielt therapieren zu können, finden auf der biologischen Seite ein mögliches Korrelat dahingehend, dass die drei aminergen Neurotransmittersysteme, die als primäres Target für praktisch alle Antidepressiva gelten können (Serotonin, Noradrenalin, Dopamin), trotz einer sehr bescheidenen Anzahl der jeweiligen Nervenzellen im Gehirn bezogen auf die Gesamtheit, sämtliche zentralnervösen Funktionen modulierend beeinflussen [67]. Alle drei Transmittersysteme sind darüber hinaus sehr komplex miteinander über aktivierende, aber auch inhibitorische Strukturen verschaltet, sodass selbst der spezifische Angriff an nur einem Target in einem Neurotransmittersystem immer auch die anderen Transmittersystem mit beeinflusst [10–12, 32]. Das heißt, in der Endstrecke ist letztlich der selektive Angriff einer solchen Substanz gar nicht mehr so selektiv, sodass es dann doch logisch erscheint, dass das depressive Syndrom in seiner Gesamtheit reagiert. Das gilt auch für Agomelatin, wo wahrscheinlich sogar der Angriff an dem Melatoninrezeptor als hochselektivem spezifischem Angriffspunkt über modulierende Effekte indirekt die drei biogenen Neurotransmitter beeinflusst und letztlich auch erklären kann, dass Agomelatin sich in der Breite der therapeutischen Wirkung auf das depressive Syndrom nicht wesentlich von den anderen Substanzen unterscheidet [18].
Auf der anderen Seite können aber auch einzelne Symptome des depressiven Syndroms über die sonstige Verbesserung hinaus persistieren und eine vollständige Remission erschweren [10, 11]. Hier kann eine auf den unterschiedlichen pharmakologischen Eigenschaften der Antidepressiva basierte Therapie sinnvoll sein [10, 13, 94]. Beispiele wären der Einsatz von dualen Wiederaufnahmehemmern wie Venlafaxin oder Duloxetin bei chronischen Schmerzsymptomen oder der Einsatz von auch H1-antagonistischen Substanzen wie Mirtazapin bei ausgeprägten Schlafproblemen.
Zu solchen über die sonstige Remission persistierenden Symptomen gehören auch kognitive Störungen [69]. Kognitive Dysfunktion ist einer der wichtigsten Teilaspekte des depressiven Syndroms; sie betrifft häufig Bereiche wie Aufmerksamkeit, Arbeitsgedächtnis und Exekutivfunktionen [10, 13, 69] und spielt damit eine wichtige Rolle in der funktionellen Beeinträchtigung depressiver Patienten. Teile der kognitiven Störungen sind eng mit dem Syndrom verbunden, treten zusammen mit den anderen Symptomen auf und klingen auch zusammen mit den anderen Symptomen im natürlichen Verlauf oder nach therapeutischer Intervention ab [64]. Ein erheblicher Teil der kognitiven Beeinträchtigung kann aber offensichtlich losgekoppelt vom eigentlichen depressiven Syndrom lange nach Remission der eigentlichen depressiven Symptomatik persistieren und wesentlich zur funktionellen Beeinträchtigung depressiver Patienten beitragen, über die die Patienten häufig klagen [13, 29, 33–35, 43, 78, 82, 87, 90]. Klinisches Erscheinungsbild, Verlauf und Charakterisierung kognitiver Dysfunktionen im Rahmen der Depression sind ausführlich im aktuellen Beitrag von Otte [69] beschrieben.
Neurobiologische Mechanismen
Die Tatsache, dass kognitive Störungen eine gewisse Selbstständigkeit im Rahmen der depressiven Symptomatik aufweisen, lässt es möglich erscheinen, dass die neurobiologischen Mechanismen, die diesen kognitiven Störungen zugrunde liegen, unabhängig von der Depression sind. Wenn man diesem auf den Grund gehen möchte, ergibt sich natürlich als primäres Problem, dass wir letztlich die Neurobiologie der Depression nicht genau kennen [17, 73, 93]. Man geht allerdings heute davon aus, dass Störungen von Mechanismen der Neuroplastizität eine wichtige Rolle spielen und über einen initialen Angriff an den drei aminergen Transmittern durch die Antidepressiva korrigiert werden. Inwieweit direkte Störungen an diesen drei Transmittersystemen vorliegen, ist umstritten. Selbst für die serotonerge Neurotransmission, für die es noch die besten Evidenzen gibt, sind die Daten begrenzt (Tab. 1) [23]. Berücksichtigt man die unklare Datenlage für die Depression selbst, ist es nicht verwunderlich, dass unsere Kenntnisse zur Neurobiologie kognitiver Störungen noch begrenzter sind [20, 55, 61, 69]. Während zwar einige Aussagen über die beteiligten neuroanatomischen Strukturen möglich sind [61], sind unsere Vorstellungen über die beteiligten neurochemischen Mechanismen eher vage [84].
Tab. 1. Neurochemische Veränderung innerhalb der serotonergen Neurotransmission bei depressiven Patienten (Zusammenstellung nach Cowen, 2008). Trotz aller Variabilität weisen die Befunde auf eine biochemische Störung innerhalb der serotonergen Neurotransmission bei depressiven Patienten hin, in den meisten Fällen als „trait“-Veränderung im Sinne einer biologischen Narbe [22, 23].
|
Veränderter serotonerger Parameter bei depressiven Patienten |
Veränderung |
Signifikanz |
|
Serotoninaufnahme in Thrombozyten |
Erniedrigt |
Sehr gut |
|
Imipramin-Bindung an Thrombozyten |
Erniedrigt |
Gut |
|
5-HT2A-Rezeptorbindung an Thrombozyten |
Erhöht |
Mäßig |
|
Plasma-Tryptophan |
Erniedrigt |
Sehr gut |
|
5-HIAA im Liquor |
Erniedrigt |
Mäßig |
|
Prolactinfreisetzung durch SSRI |
Erniedrigt |
Sehr gut |
|
5-HT1A-Rezeptordichte im Gehirn (PET) |
Erniedrigt |
Sehr gut |
|
5-HT2A-Rezeptorbindung im Gehirn |
Variabel |
Schlecht |
5-HIAA: 5-Hydroxyindolylessigsäure, primärer Serotoninmetabolit
Diese in der Übersicht von Otte [69] zusammengefassten Mechanismen zeigen daher auch eine große Bandbreite, und wie vom Autor folgerichtig diskutiert, gibt es für keinen dieser Mechanismen eine so gute Evidenz, dass man ihn uneingeschränkt als Erklärung für die persistierenden kognitiven Beeinträchtigungen akzeptieren könnte. Diese Hypothesen gehen von Störungen in den für kognitive Funktionsabläufe wichtigen präfrontal-kortikalen Netzwerken aus, wie sie von verschiedenen fMRT-Untersuchungen gezeigt wurden. Auch der Hippocampus wird als betroffenes Areal möglicherweise auch im Zusammenhang mit Störungen der Cortisolachse diskutiert, die aber auch über den Hippocampus hinaus in diese Störungsbilder involviert sein könnten [77]. Andere Hypothesen gehen von Störungen im Bereich der neurotrophen Faktoren aus, und auch inflammatorische Mechanismen, die ja ebenfalls im Rahmen der Depression selbst diskutiert werden, sind als Ursache kognitiver Defizite in Betracht gezogen worden. Auffällig erscheint zunächst, dass Veränderungen der noradrenergen und/oder dopaminergen Neurotransmission eher nicht als Ursache kognitiver Störungen im Rahmen der Depression bekannt sind, obwohl beide Systeme in der Regulation von Kognition eine unbestritten wichtige Rolle spielen [10–12, 20, 61, 81] und eine pharmakologische Manipulation beider Systeme durchaus einen positiven Effekt auf Störungen der Kognition haben kann (siehe Duloxetin im Abschnitt „Effekte von Antidepressiva auf kognitive Störungen depressiver Patienten“).
Tryptophandepletion (TPD) als Indikator für eine serotonerge Narbe
Etwas anders ist die Situation bei der serotonergen Neurotransmission, die nicht nur eine wichtige allgemeine Rolle in der Regulation von Kognition spielt [20, 23]; darüber hinaus wird eine serotonerge Störung bzw. serotonerge Narbe als ätiopathogenetisch wichtiger Mechanismus kognitiver Veränderungen bei depressiven Erkrankungen, besonders im Hinblick auf die persistierenden Störungsbilder, angenommen. Diese Daten gehen hauptsächlich auf Untersuchungen mit dem Tryptophandepletionstest zurück, wo durch Gabe eines Tryptophan-freien Aminosäuregemischs die Tryptophanaufnahme ins Gehirn und damit die zentrale Serotoninsynthese deutlich reduziert ist. In Untersuchungen an Tieren [3, 42], an gesunden Probanden [57] sowie depressiven Patienten bzw. Personen mit einem Depressionsrisiko (remittierte Patienten, Verwandte ersten Grades von depressiven Patienten) konnte gezeigt werden, dass durch Tryptophandepletion (TPD) Kognitionsverschlechterungen ausgelöst werden konnten [6, 22, 80]. Bei gesunden Probanden, siehe die ausführliche Übersicht von Mendelsohn et al. [57], kann durch Tryptophandepletion eine kognitive Verschlechterung ohne Veränderungen der Stimmungslage ausgelöst werden. Im Gegensatz dazu kann TPD bei Gesunden mit einer biologischen Narbe auch Stimmungsverschlechterungen auslösen, so beispielsweise bei unterschiedlichen Serotonintransporter(SERT)-Polymorphismen [83] oder bei remittierten depressiven Patienten, bei denen aber die Effekte der TPD auf die Kognition oft deutlicher sind als die Effekte auf die Stimmungslage [6, 23, 59, 74, 79, 80]. Ein Beispiel für die Kognitionsverschlechterung nach TPD ist die Arbeit von Porter et al. [74], wo 16 remittierte depressive Patienten und 17 Gesunde nach TPD mit verschiedenen kognitiven Tests untersucht wurden, wie in Abbildung 2 am Beispiel des modifizierten MMSE gezeigt. Der Befund (siehe Abb. 2), dass remittierte depressive Patienten nach TPD im Bereich der Kognition empfindlicher als Gesunde, mit einer gewissen Kognitionsverschlechterung reagieren, ist mehrfach repliziert worden [22, 31, 36]. Auf der anderen Seite kann TPD auch positive Effekte auf die Kognition haben, besonders bei Gesunden [57], was parallel geht mit den pharmakologischen Effekten von SSRI auf die Kognition Gesunder, wo positive aber auch negative Effekte gesehen wurden, wie im Folgenden noch kurz diskutiert wird. Damit ist die Beziehung zwischen Serotonin und Kognition im Einzelfall etwas komplizierter als die sicher zutreffende, aber eben vereinfachende Aussage, dass erniedrigtes Serotonin mit einer beeinträchtigten Kognition einhergeht [22].
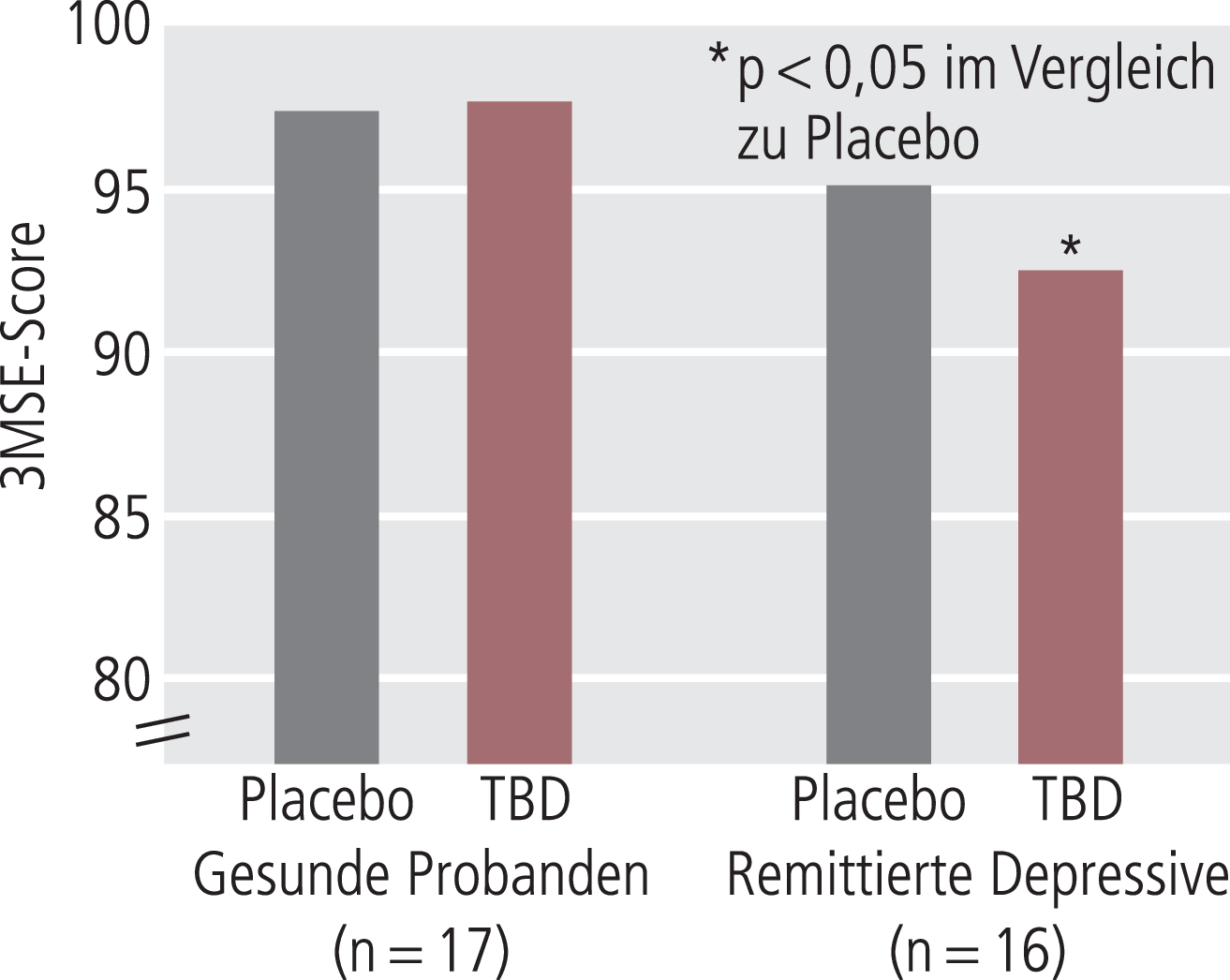
Abb. 2. Effekt einer Tryptophandepletion (TPD) auf die kognitive Funktion bei Gesunden und bei remittierten Depressions-Patienten. Bei Gesunden hatte TDP keinen Effekt auf den modifizierten MMSE (3MSE), reduzierte aber bei den remittierten Patienten den 3MSE-Score signifikant. Die Daten stammen aus der Arbeit von Porter et al. [74]
In Übereinstimmung mit der wichtigen Rolle von Serotonin zusammen mit den beiden anderen biogenen Aminen Dopamin und Noradrenalin [81] führt TPD auch bei Tieren zu einer Verschlechterung bestimmter kognitiver Funktionen [42, 44, 49]. Gerade wegen der komplexen Verschaltung der drei biogenen Amine sind die Befunde von Ardis et al. [3] wichtig, dass durch TPD an der Ratte die Noradrenalin- und Dopamin-Spiegel im Gehirn weitgehend unverändert bleiben.
Unter den vielen experimentellen Modellen, die zur Verfügung stehen, um im Tierexperiment kognitive Funktionen zu erfassen, hat sich der Objekt-Wiedererkennungstest [2] besonders bewährt, um kognitive Defizite nach TPD abzubilden [45, 68], wie am Beispiel von unbehandelten Ratten in Abbildung 3 gezeigt. Wichtig ist hier, dass der Effekt von TPD auf die Kognition bei vorliegender biologischer Narbe, beispielsweise einer Reduktion der Serotonintransporter(SERT)-Dichte, ausgeprägter ist (Abb. 4) [68]. Dies zeigt eine gewisse Parallele zu den Befunden am Menschen, dass SERT-Polymorphismen die durch TPD hervorgerufene Kognitionsverschlechterung beeinflussen [83]. Ohne diese Befunde überbewerten zu wollen, geben sie doch einen Hinweis, dass die Sonderstellung kognitiver Beeinträchtigungen bei depressiven Patienten – im Hinblick auf eine gewisse Unabhängigkeit von der Restsymptomatik und auf die Persistenz über die Remission hinaus – mit serotonergen Mechanismen im Sinne einer biologischen Narbe assoziiert zu sein scheint. Die Befunde sind zwar komplex, können aber in einer vereinfachten Darstellung dahingehend zusammengefasst werden, dass Reduktion der extrazellulären Serotoninkonzentration mit gestörter Kognition parallel geht, wobei besonders Gedächtniskonsolidierung betroffen ist [23]. Einschränkend muss man aber auch darauf hinweisen, dass durch die komplexe Beteiligung verschiedener Serotoninrezeptortypen mit unterschiedlicher Lokalisierung (postsynaptisch eher aktivierend bzw. präsynaptisch eher hemmend) im Einzelfall auch weniger Serotonin mit positiven bzw. mehr Serotonin mit negativen Effekten auf die Kognition einhergehen kann [20, 23].
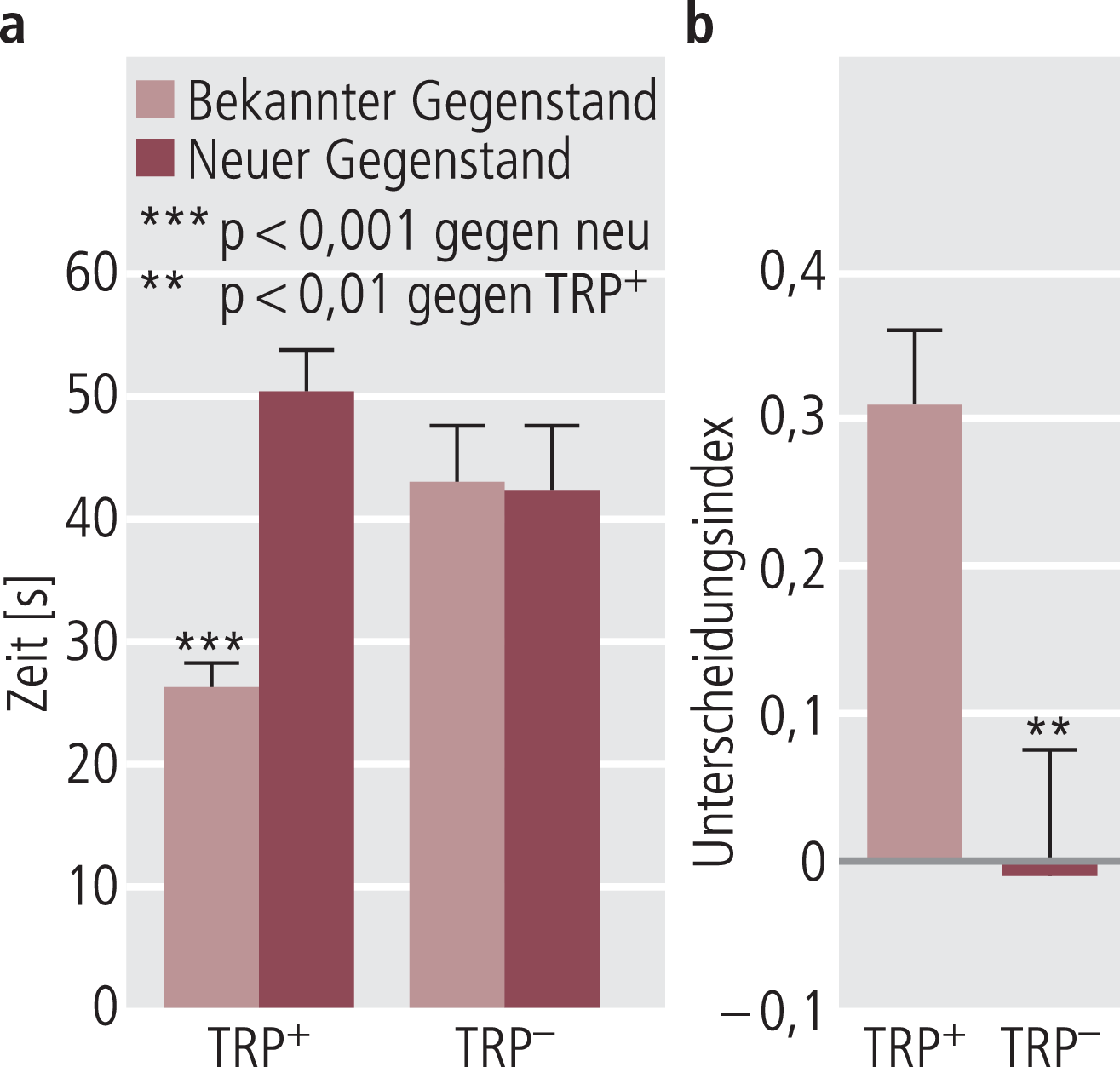
Abb. 3. Eine Serotoninreduktion durch 14-tägige Tryptophan-freie Diät führt zu einer Störung des Erinnerungsvermögens von Ratten im Gegenstands-Wiedererkennungstest. a) Im Vergleich zu den Kontrollen (Tryptophan-haltige Diät, TRP+), die sich deutlich länger mit dem neuen Gegenstand beschäftigen, führt die Serotoninreduktion durch die Tryptophan-freie Diät (TRP–) zu einer schlechteren Erinnerung in den bekannten Gegenstand, damit zu einer längeren Beschäftigung mit dem eigentlichen gekannten Gegenstand. b) Dies reduziert auch den Unterscheidungsindex auf praktisch Null. Die Daten stammen aus der Arbeit von Jenkins et al. [45].
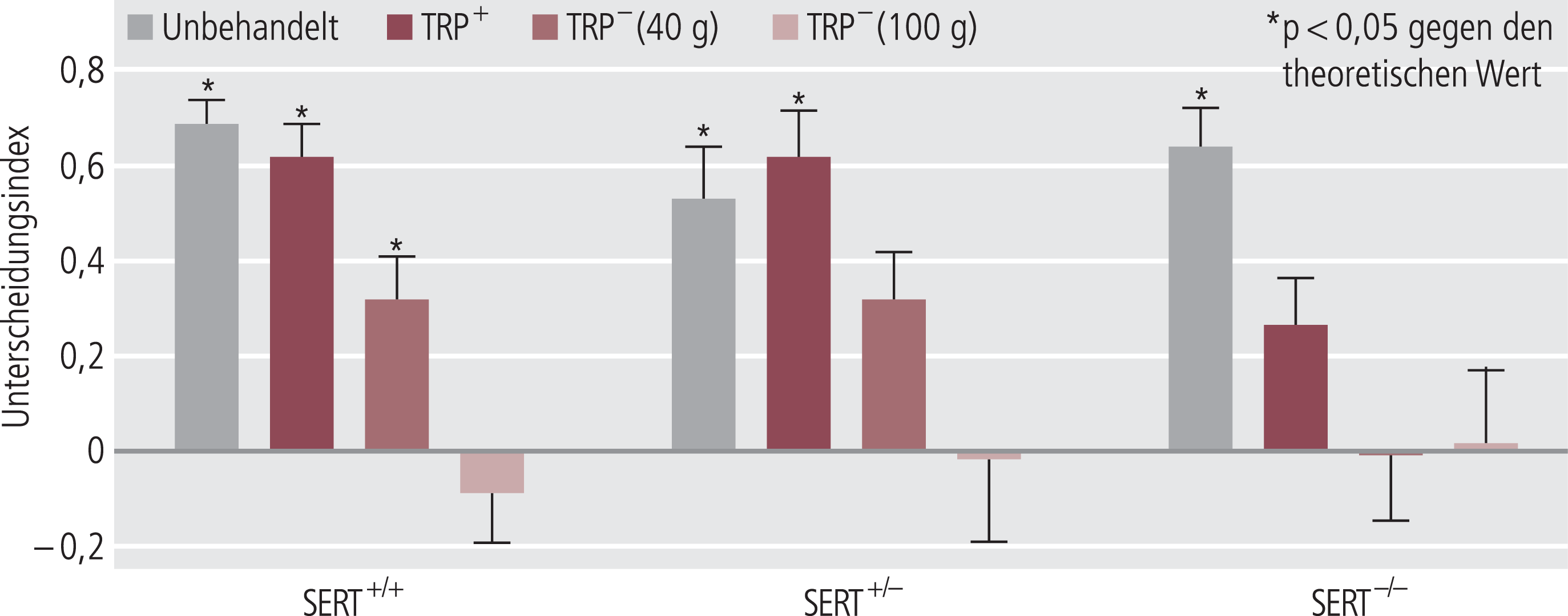
Abb. 4. Eine serotonerge Narbe (Abnahme der SERT-[Serotonintransporter-]Expression) verstärkt den Effekt einer Tryptophandepletion (TDP) auf die Kognition im Tierversuch. Im Gegenstand-Erinnerungstest wird eine Kognitionsverschlechterung über die Zeit ermittelt, mit der sich die Versuchstiere mit einem neuen, im Vergleich zu einem bekannten Gegenstand beschäftigen. Der theoretische Wert von 1,0 würde 100%ige Beschäftigung mit dem neuen Gegenstand bedeuten, also vollständige Erinnerung des bekannten Gegenstandes. Gesunde Tiere (graue Säule) haben hier einen Wert von etwa 0,7 (d.h., sie beschäftigen sich auch noch etwas mit dem bekannten Objekt). Bei zunehmender Abnahme der SERT-Expression (SERT+/– bzw. SERT–/–) ändert sich die kognitive Leistungsfähigkeit bei den Kontrolltieren (graue Säulen) nur geringfügig. TDP hatte dagegen dosisabhängig einen negativen Effekt auf das Gedächtnis (TRP– [40 g bzw. 100 g Tryptophan-freies Aminosäuregemisch pro 100 ml der oralen Behandlungslösung, gegeben als 2×10 ml innerhalb der zwei Stunden vor dem Versuch]). Ein Tryptophan-haltiges Aminosäuregemisch (dunkelrote Säulen, reduziert Tryptophan-haltig) zeigte einen geringeren Effekt. Alle Effekte waren bei den SERT–/–Tieren am deutlichsten ausgeprägt. Die Daten stammen aus der Arbeit von Olivier et al. [68].
Die eben angesprochene Komplexität des Einflusses von Serotonin auf kognitive Funktionen mit negativen, manchmal aber auch positiven Effekten einer experimentellen Serotoninverarmung durch TPD spiegelt sich auch in den Wirkungen von SSRI auf die Kognition gesunder Probanden wider, wo eine Erhöhung der extrazellulären Serotoninkonzentration erreicht wird. Die vereinfachte, aber doch durch viele experimentelle Befunde getragene Aussage, dass Serotoninmangel eher mit kognitiven Defiziten assoziiert ist, könnte zu der Schlussfolgerung führen, dass eine Erhöhung der extrazellulären Serotoninkonzentration durch SSRI die Kognition verbessern müsste. Dies ist allerdings nicht immer der Fall und eine Reihe von Befunden zeigt, dass bei gesunden Probanden SSRI sehr variable Effekte auf kognitive Parameter haben, die sich von positiv im Sinne einer Verbesserung bis häufig auch negativ im Sinne einer Verschlechterung darstellen können (Tab. 2). Eine pharmakologische Serotoninerhöhung allein erscheint damit nicht ausreichend, um eine sichere Kognitionsverbesserung zu erreichen.
Tab. 2. Effekte von SSRI auf kognitive Funktionsparameter gesunder Probanden [Zusammenstellung nach 22]
|
Name des SSRI |
Wirkung auf Kognition |
|
Citalopram |
|
|
Escitalopram |
|
|
Fluvoxamin |
|
|
Sertralin |
|
Effekte von Antidepressiva auf kognitive Störungen depressiver Patienten
Eine spezifische Verbesserung kognitiver Beeinträchtigung depressiver Patienten war und ist damit ein bekanntes, aber eher ungelöstes therapeutisches Problem [69], besonders auch im Hinblick auf unsere heutigen Kenntnisse einer gewissen eigenständigen Rolle kognitiver Symptome innerhalb des depressiven Syndroms und als wichtiges Residualsymptom mit erheblichen Funktionsbeeinträchtigungen nach sonstiger Remission [22, 54, 55]. Es ist bekannt, dass im Rahmen der Verbesserung des depressiven Syndroms unter antidepressiver Therapie auch kognitive Symptome ansprechen und eine parallele Verbesserung zeigen [89]. Auf der anderen Seite gibt es aber nur wenige Untersuchungen, die sich mit der Fragestellung beschäftigt haben, ob kognitive Störungen depressiver Patienten auch über die Verbesserung im Rahmen des depressiven Syndroms therapeutisch ansprechen [33–35, 54, 55].
Die damit verbundene Frage, ob unterschiedliche Antidepressiva im Hinblick auf eine Verbesserung kognitiver Dysfunktion depressiver Patienten über die Verbesserung des depressiven Syndroms hinaus wirksam sind, ist in verschiedenen Übersichtsarbeiten diskutiert worden [1, 12, 89, 91]. Um eine sehr komplexe Datenlage möglichst knapp zusammenzufassen, kann man davon ausgehen, dass analog zu den sehr variablen Befunden der spezifische Serotonin-Wiederaufnahmehemmer bei gesunden Probanden keine belastbaren Befunde vorhanden sind, die eine spezifische Verbesserung kognitiver Symptome durch Antidepressiva im Allgemeinen oder durch SSRI im Speziellen zeigen. Dies ist zunächst für die älteren, meist auch anticholinerg wirkenden Antidepressiva nicht verwunderlich, aber für die SSRI eingedenk einer häufig vorhandenen serotonergen Narbe depressiver Patienten eher unerwartet. Der Widerspruch ist offenkundig, da auf der einen Seite ein niedriger Serotoninspiegel einen relativ reproduzierbaren negativen Effekt auf kognitive Parameter hat (siehe Abschnitt „Tryptophandepletion [TPD] als Indikator für eine serotonerge Narbe“) und man von Substanzen, die den Serotoninspiegel erhöhen, deutlichere und spezifischere Effekte erwarten würde. Die Erklärung liegt, wie bereits andiskutiert, in der Problematik, dass Serotonin über die verschiedenen Serotoninrezeptoren sowohl positive wie auch negative Effekte auf kognitive Parameter haben kann, die offensichtlich durch die unspezifische Anhebung der Serotoninkonzentration durch SSRI alle aktiviert werden können, was zu variablen Effekten auf die Kognition führen kann.
Antidepressiva, die im Gegensatz zu den SSRI etwas deutlichere Depressionssyndrom-unabhängige positive Effekte auf kognitive Defizite gezeigt haben, sind Bupropion und duale Serotonin- und Noradrenalin-Wiederaufnahmehemmer besonders Duloxetin. Für Bupropion sind die Daten eher anekdotisch [40] und wegen fehlender Verblindung auch nicht sehr aussagekräftig (Gruppengröße 37 Patienten). Für den spezifischen Serotonin- und Noradrenalin-Wiederaufnahmehemmer Duloxetin gibt es dagegen einige kontrollierte Untersuchungen, die auf eine spezifische Wirkung auf kognitive Symptome depressiver Patienten hinweisen. In einer doppelblinden Vergleichsstudie mit Escitalopram gegen Duloxetin an depressiven Patienten über 24 Wochen (Gruppengröße 37 Patienten) hatten beide Substanzen einen ähnlichen Effekt auf die depressive Symptomatik; beide verbesserten auch die kognitiven Symptome, wobei allerdings Duloxetin überlegen war, besonders im Bereich von episodischem Gedächtnis und Arbeitsgedächtnis. Diese Überlegenheit blieb auch nach Beendigung der Therapie in der Remissionsphase stabil [38, 39, 41]. Sehr viel aussagekräftiger ist die klassische Untersuchung von Raskin et al. [76], wo Duloxetin in einer Vergleichsuntersuchung gegen Placebo bei depressiven Patienten kognitive Defizite (im Bereich verbalen Lernens) deutlich stärker als Placebo verbesserte und wo der Effekt auf die Kognition teilweise unabhängig von Depressions-verbessernden Effekten der Substanz war, was mittels Pfadanalyse gezeigt wurde. In einer aktuellen offenen Studie konnte gezeigt werden, dass sich der positive Effekt von Duloxetin über verschiedene kognitive Domänen erstreckte wie sichtbares oder verbales Lernen und Gedächtnis, affektive Entscheidungsfindung und Reaktionskontrolle und etwas geringer auch im Bereich der Exekutivfunktionen [28]. Auch hier wurde bestätigt, dass die positiven Effekte auf Kognition unabhängig von den Depressions-lösenden Effekten sind. Dass die Noradrenalin-Wiederaufnahme-hemmende Komponente von Duloxetin hier eine wichtige Rolle zu spielen scheint, geht auch aus einer Pilotuntersuchung hervor, wo gezeigt werden konnte, dass die Zusatzbehandlung depressiver Patienten, die unter Escitalopram-Therapie standen, mit dem Noradrenalin-Wiederaufnahmehemmer Atomoxetin zu Verbesserungen kognitiver Beeinträchtigungen führte [7]. Des Weiteren hatte der Noradrenalin-Wiederaufnahmehemmer Reboxetin in einer Doppelblinduntersuchung positive Effekte auf gestörte kognitive Parameter depressiver Patienten, die Vergleichssubstanz Paroxetin war hier nicht wirksam. Verbesserungen von kognitiven Parametern korrelierten nicht mit Verbesserungen der Depressivität [25].
Dass Noradrenalin-Wiederaufnahmehemmung mit Kognition-verbessernden Effekten einhergehen kann, ist nicht unerwartet, wenn man den therapeutischen Einsatz von Atomoxetin beim Aufmerksamkeitsdefizit-/Hyperaktivitäts-Syndrom (ADHS) betrachtet. Es ist sicher mechanistisch auch damit verbunden, dass im präfrontalen Kortex, einer wichtigen Regulationsstelle für kognitive Parameter wie Aufmerksamkeit und Arbeitsgedächtnis, wegen des Fehlens von Dopamintransportern Noradrenalin-Wiederaufnahmehemmer auch die neuronale Rückaufnahme von Dopamin hemmen, was in dieser Struktur eine sehr wichtige Rolle für die Kognition spielt [15, 88]. Einschränkend muss man allerdings konstatieren, dass für beide klinisch eingesetzte Substanzen (Reboxetin und Duloxetin) zwar in bestimmten Anordnungen positive Effekte gesehen wurden, die allerdings relativ bescheiden waren, und dass sich generell beide Substanzen im Spektrum der klinisch eingesetzten Antidepressiva als Kognitionsverbesserer nicht besonders platzieren konnten.
Effekte von Vortioxetin
Eine pharmakologisch ganz andere Substanz, die besondere Effekte auf kognitive Parameter gezeigt hat, ist Vortioxetin. Es hemmt den Serotonintransporter und zeigt gleichzeitig noch agonistische, partialagonistische und antagonistische Effekte an einer Reihe von Serotoninrezeptoren (Abb. 5) [85]. In einer großen doppelblinden Placebo-kontrollierten Studie an depressiven Patienten, wo ähnliche Instrumente eingesetzt wurden wie in der bereits zitierten Studie mit Duloxetin von Raskin et al. [76], konnten für Vortioxetin deutliche Effekte auf kognitive Parameter in einer primär festgelegten Sekundäranalyse gezeigt werden (Abb. 6) [47]. Vortioxetin hatte dabei eine etwas breitere Wirkung auf neuropsychologische kognitive Parameter, als dies für Duloxetin beobachtet wurde: Es zeigte nicht nur eine signifikante Verbesserung im RAVLT (Rey auditory verbal learning test), in dem auch, wie schon in der Untersuchung von Raskin et al. [76] gezeigt, Duloxetin wirksam war, sondern war auch signifikant wirksam (gegen Placebo) im Bereich Geschwindigkeit, Exekutivfunktionen und Aufmerksamkeit (DSST-Test, Digit symbol substitution test), wo Duloxetin nicht signifikant wirksam war (Abb. 6), wieder in Bestätigung der Befunde von Raskin et al. [76].
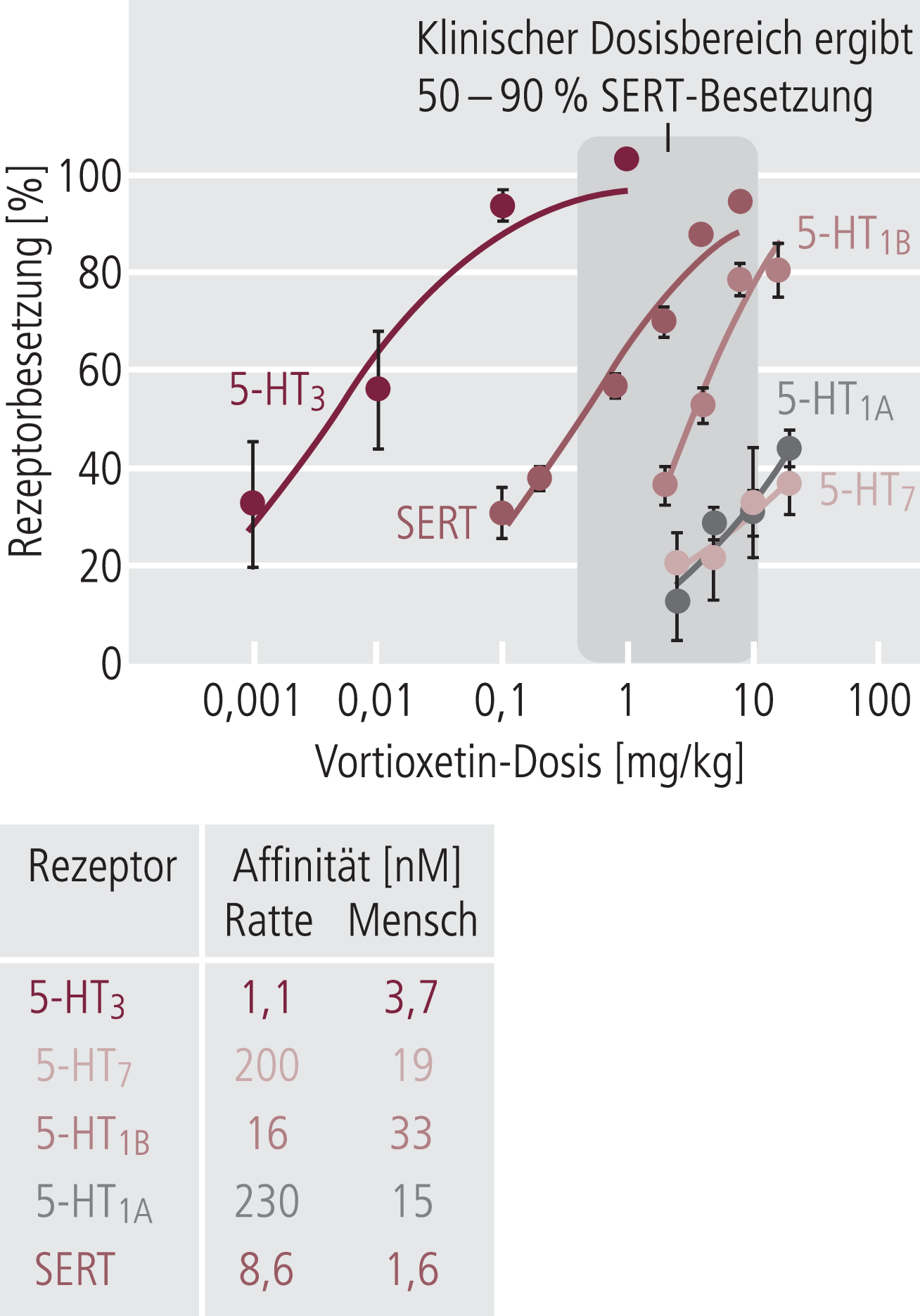
Abb. 5. Pharmakologie des „multi-modalen“ Antidepressivums Vortioxetin. Als Serotonin-Wiederaufnahmehemmer bindet es im therapeutischen Bereich an den Serotonintransporter (SERT), daneben aber auch sehr stark an den 5-HT3-Rezeptor und mit etwas geringeren Affinitäten an die 5-HT1A-, 5-HT1B- und 5-HT7-Rezeptoren.
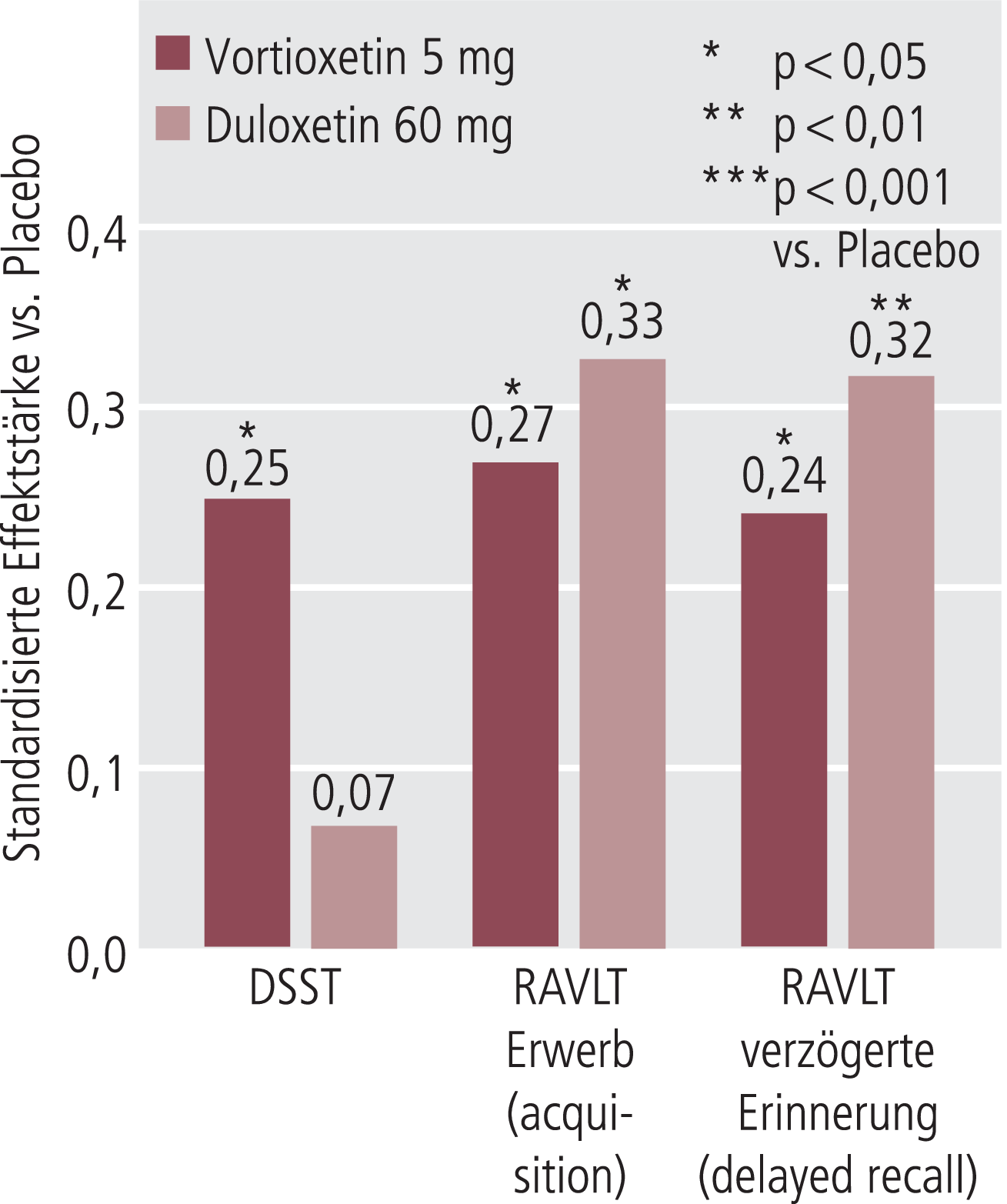
Abb. 6. Kognitionsverbesserung bei älteren depressiven Patienten unabhängig vom antidepressiven Effekt. In der Arbeit von Katona et al. [47] wurde im Rahmen einer Depressionsstudie der Effekt von Duloxetin (60 mg/Tag) und Vortioxetin (5 mg/Tag) auf Kognition überprüft. Die a priori als Sekundärkriterien gewählten Tests (DSST und RAVLT) wurden aus der Arbeit von Raskin et al. [76] übernommen. DSST: Digit symbol substitution test; RAVLT: Rey auditory verbal learning test
Auch hier waren die kognitiven Verbesserungen unter Vortioxetin gemäß Pfadanalyse weitgehend unabhängig von dem parallel gesehenen Effekt dieser Substanz auf die depressive Symptomatik [47]. Diese Daten wurden in einer kürzlich publizierten Doppelblind-Untersuchung an depressiven Patienten bestätigt, wo Kognitions-verbessernde Effekte von Vortioxetin im Vergleich zu Placebo als a priori festgelegter primärer Wirksamkeitsnachweis untersucht wurden und wo gezeigt werden konnte, dass Vortioxetin in einer ganzen Reihe von unterschiedlichen kognitiven Domänen positive Effekte zeigte [56]. In einer weiteren Doppelblinduntersuchung an erwachsenen Patienten mit einer majoren Depression und subjektiv beklagten kognitiven Dysfunktionen (primärer Endpunkt) wurde Vortioxetin gegenüber Placebo untersucht. Duloxetin fungierte als Aktivreferenz. Wie erwartet, zeigte nur Vortioxetin im DSST eine signifikante Verbesserung gegenüber Placebo, also im wichtigen Bereich Geschwindigkeit, Exekutivfunktionen und Aufmerksamkeit, nicht aber Duloxetin [53]. Eine Pfadanalyse deutet auch hier eher auf einen direkten Effekt von Vortioxetin auf die kognitive Performanz und die funktionelle Leistungsfähigkeit, weniger auf einen indirekten Effekt über die Verbesserung depressiver Symptome.
Diese spezifischen Effekte von Vortioxetin werfen die Frage nach dem Wirkungsmechanismus auf. War die bessere kognitive Wirksamkeit von Duloxetin gegenüber SSRI noch durch die zusätzliche Beeinflussung noradrenerger und dopaminerger Mechanismen relativ leicht erklärbar, haben wir im Fall von Vortioxetin eine Substanz, die nur primär im serotonergen System angreift, allerdings über verschiedene Mechanismen, von der Hemmung von SERT über agonistische, partialagonistische und antagonistische Effekte an verschiedenen Serotoninrezeptoren: Im Folgenden daher eine Zusammenfassung zum aktuellen Stand der kognitiven Pharmakologie von Vortioxetin.
Pharmakologische Eigenschaften von Vortioxetin
Effekte auf die Kognition: Vortioxetin versus andere Antidepressiva
Die Darstellung von kognitionsverbessernden Effekten im Tierversuch ist mit dem Problem behaftet, dass im Tiermodell kognitive Parameter nur indirekt über Verhaltensänderungen erfasst werden können, was natürlich einen großen Verlust an Empfindlichkeit bedeutet. Die Erfassung kognitionsverbessernder Effekte von Arzneimitteln im Allgemeinen und von Antidepressiva im Speziellen ist daher im Tierversuch keinesfalls generell leichter als im kontrollierten Test am Menschen. Bedenkt man jetzt die großen Probleme, die bei der Erfassung positiver Effekte von Antidepressiva am Menschen existieren, ist es nachvollziehbar, dass die Daten für Antidepressiva im Tierversuch sehr widersprüchlich sind, wie in einigen Übersichten zusammengefasst [63, 72]. Wenn man versucht, diese sehr komplexe Datenlage auf einen einfachen Nenner herunterzubrechen, muss man feststellen, dass es praktisch für keines der gängigen Antidepressiva möglich war, einen durchgehend positiven Effekt auf kognitive Funktionen im Tierversuch sicher zu belegen. Auch hier ist allerdings die Datenlage für Duloxetin etwas besser [72].
Wie zuvor dargestellt, haben besonders der duale Wiederaufnahmehemmer Duloxetin und die neue Substanz Vortioxetin klarere Effekte auf die Kognition von depressiven Patienten gezeigt als die meisten anderen Antidepressiva. Während sich der Vorteil von Duloxetin leicht über die Erhöhung von Noradrenalin und Dopamin (präfrontal) erklären lässt, besonders vor dem Hintergrund der wichtigen Rolle, die beide Systeme für die Kognition spielen [11], ist die Erklärung für die besonders gute Wirkung von Vortioxetin auf kognitive Störungen depressiver Patienten (siehe Absatz 1.4) deutlich komplexer. Während es bei Duloxetin im Fall einer anderen dualen Substanz (Venlafaxin) einige analoge Effekte gibt [16, 63, 72], betritt man mit Vortioxetin Neuland.
Im ersten Bericht zu diesem Thema zeigten Mork et al. [66], dass Vortioxetin dosisabhängig die Erinnerung an eine unangenehme, mit einem Stromstoß verbundene Situation verbesserte (Contexual fear conditioning); zu dieser Wirkung könnten noch anxiolytische Effekte der Substanz beitragen [30, 85]. Der Nachteil dieses Tests, dass das Gedächtnis über eine unangenehme Situation als relativ belastendes Ereignis abgegriffen wird, wird in dem heute gerne verwendeten „object recognition test“ umgangen, in dem die natürliche Neugier der Tiere gegenüber einem neuen Gegenstand im Käfig ausgenutzt wird [2]. Je kürzer die Zeit ist, mit der sie sich beim zweiten Kontakt noch mit dem bekannten Gegenstand beschäftigen, desto besser ist die Erinnerung an den Erstkontakt, wie im Experiment durch Zugabe eines bisher ungekannten Gegenstands bestimmt wird (Abb. 3 und 4). Auch in diesem Test, dem natürliches, nicht trainiertes Verhalten, zugrunde liegt, zeigte Vortioxetin Gedächtnis-verbessernde Effekte, ähnlich wie die hier benutzte aktive Kontrolle (der Acetylcholinesterasehemmer Donepezil) [66] (Abb. 7).
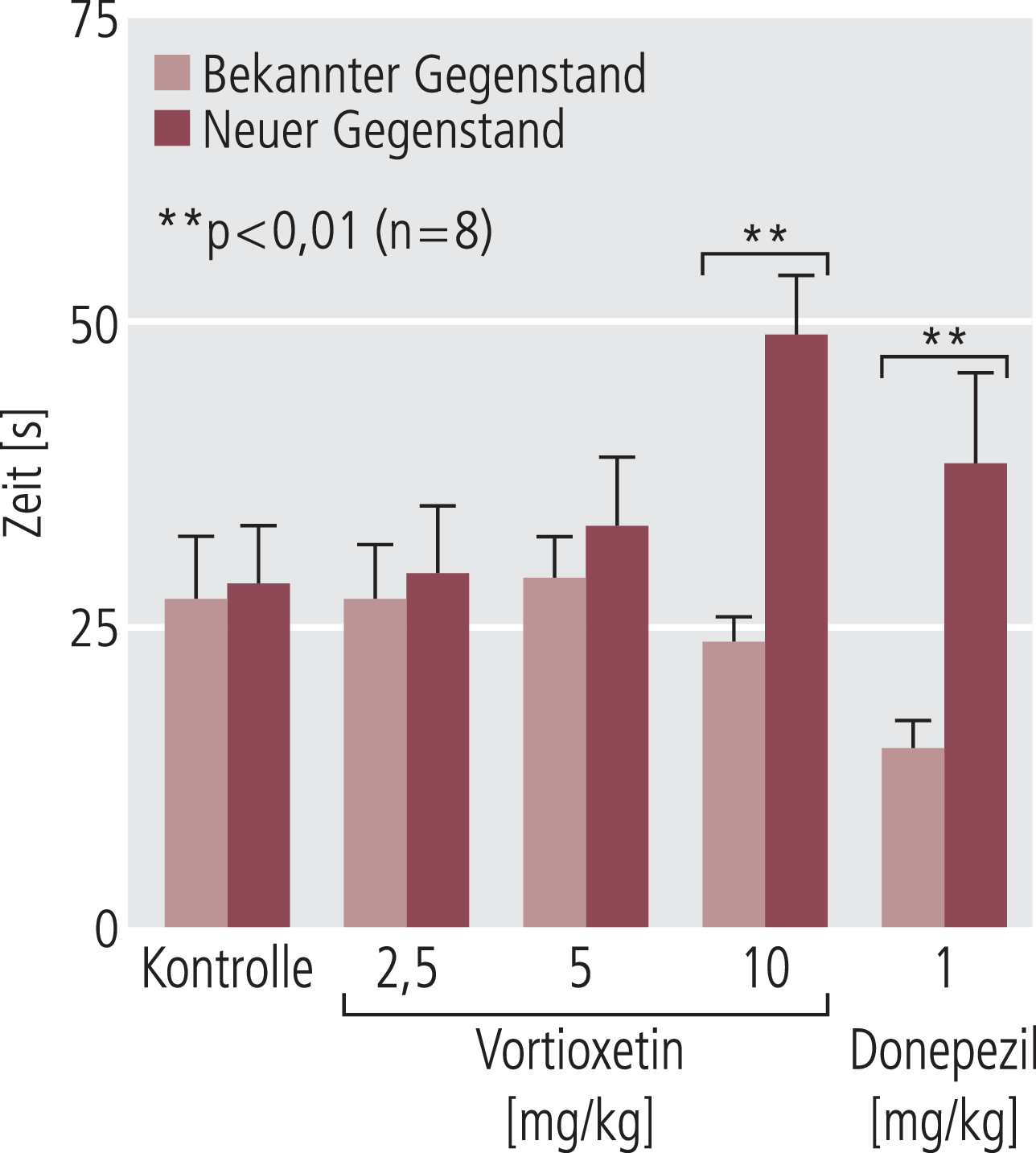
Abb. 7. Vortioxetin und Donepezil (als kognitionsverbessernde Vergleichssubstanz) sind ähnlich im Gegenstand-Wiedererkennungs-Test wirksam (längere Zeit Beschäftigung mit dem neuen Gegenstand). Die Daten sind der Arbeit von Jenkins et al. [45] entnommen
Beide Tests wurden an gesunden Ratten durchgeführt und sind daher begrenzt übertragbar auf die kognitiven Dysfunktionen depressiver Patienten. Durch vorgeschaltete Tryptophandepletion konnte dieser Test aber etwas näher an die pathophysiologische Situation depressiver Patienten herangeführt werden. Wie im Kapitel „Neurobiologische Mechanismen“ ausgeführt, kann man zwar schon an gesunden Probanden durch Tryptophandepletion eine Verschlechterung kognitiver Funktionen auslösen, die aber bei remittierten depressiven Patienten deutlich intensiver ausgeprägt sind. Analog zu diesen Befunden am Patienten bewirkt TPD bei Tieren mit einer biologischen Narbe innerhalb der serotonergen Neurotransmission (partieller oder totaler SERT-Knock-out) eine noch deutlichere Verschlechterung der Kognition (Abb. 4) [68]. Vortioxetin zeigt in diesem Modell der experimentellen Störung der serotonergen Neurotransmission mit gewisser Analogie zur Situation am Menschen eine sehr deutliche Verbesserung der Kognition, die schon bei wesentlich niedrigeren Dosen gesehen wurde als die Effekte bei den gesunden Tieren (Abb. 8) [24]. Ebenfalls in gewisser Parallelität zu den spezifischen Effekten von Vortioxetin auf die Kognition depressiver Patienten war in diesem Modell der SSRI Escitalopram nicht wirksam, während Duloxetin gewisse Effekte zeigte, die aber geringer als die von Vortioxetin waren (Abb. 9) [46].
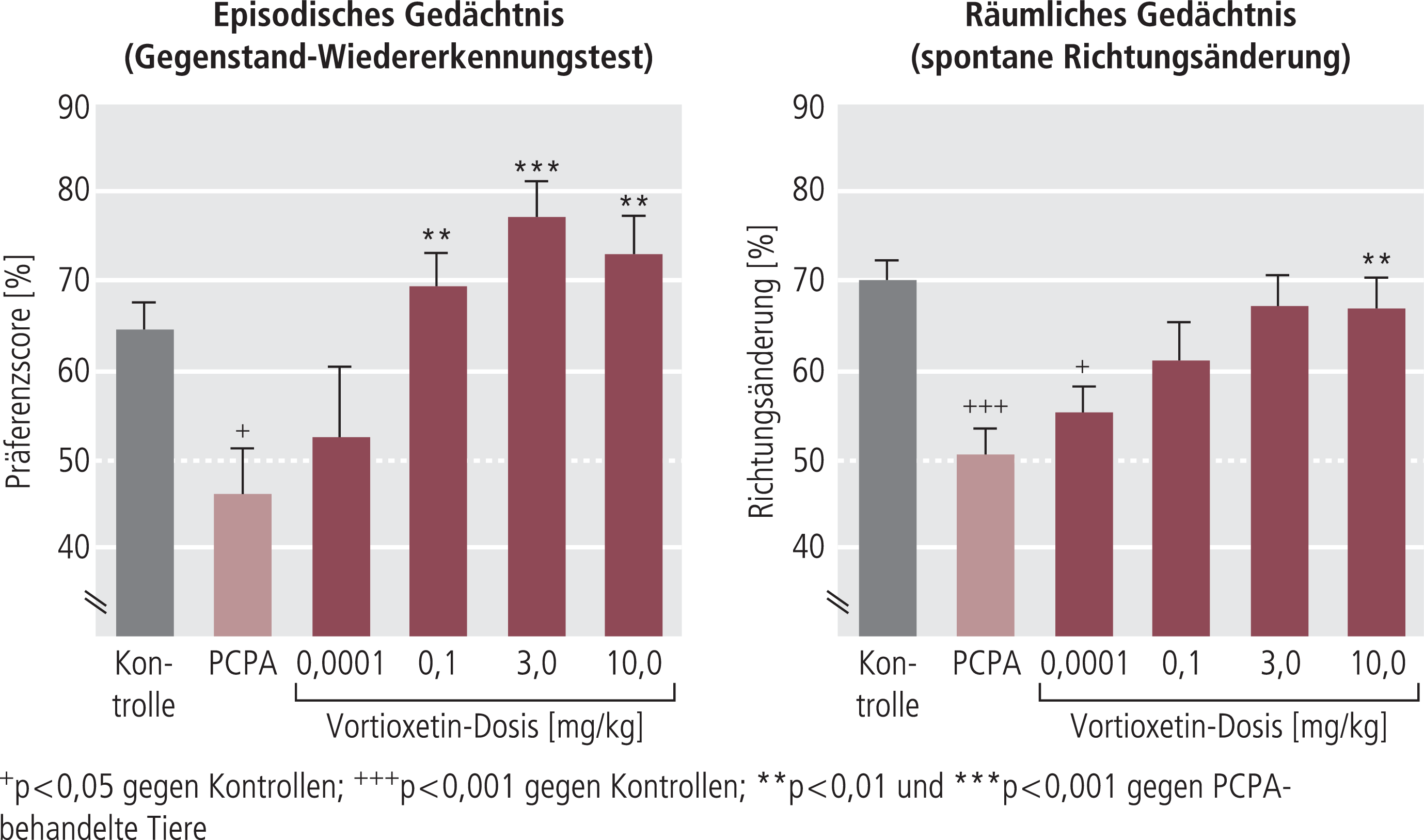
Abb. 8. Vortioxetin verbessert durch Tryptophandepletion (TPD) ausgelöste kognitive Dysfunktion. TPD (im Tierversuch üblicherweise durchgeführt durch Gabe eines Hemmstoffs der Serotoninsynthese wie hier Parachlorphenylalanin, PCPA) führt im Objekt-Wiedererkennungs-Test (siehe Abb. 2) bzw. im Test der spontanen Richtungsänderung zur Verschlechterung des Erinnerungsvermögens. Im Test der spontanen Richtungsänderung haben die Tiere in einer T-Strecke nach rechts oder links zu gehen in der Hoffung, eine Futterbelohnung zu finden. Wenn sie im akuten Test rechts nichts gefunden haben, gehen sie im anschließenden Test nach links, um ihre Chance auf eine Futterbelohnung zu bewahren. Gesunde Tiere alternieren fast 100%ig, Tiere mit einer serotonergen Narbe (TPD) kaum, sie wählen ihren Weg zufällig. In beiden Versuchen wurde die Verschlechterung des Erinnerungsvermögens durch TPD von Vortioxetin praktisch aufgehoben. Die Daten stammen aus der Arbeit von Jensen et al. [46].
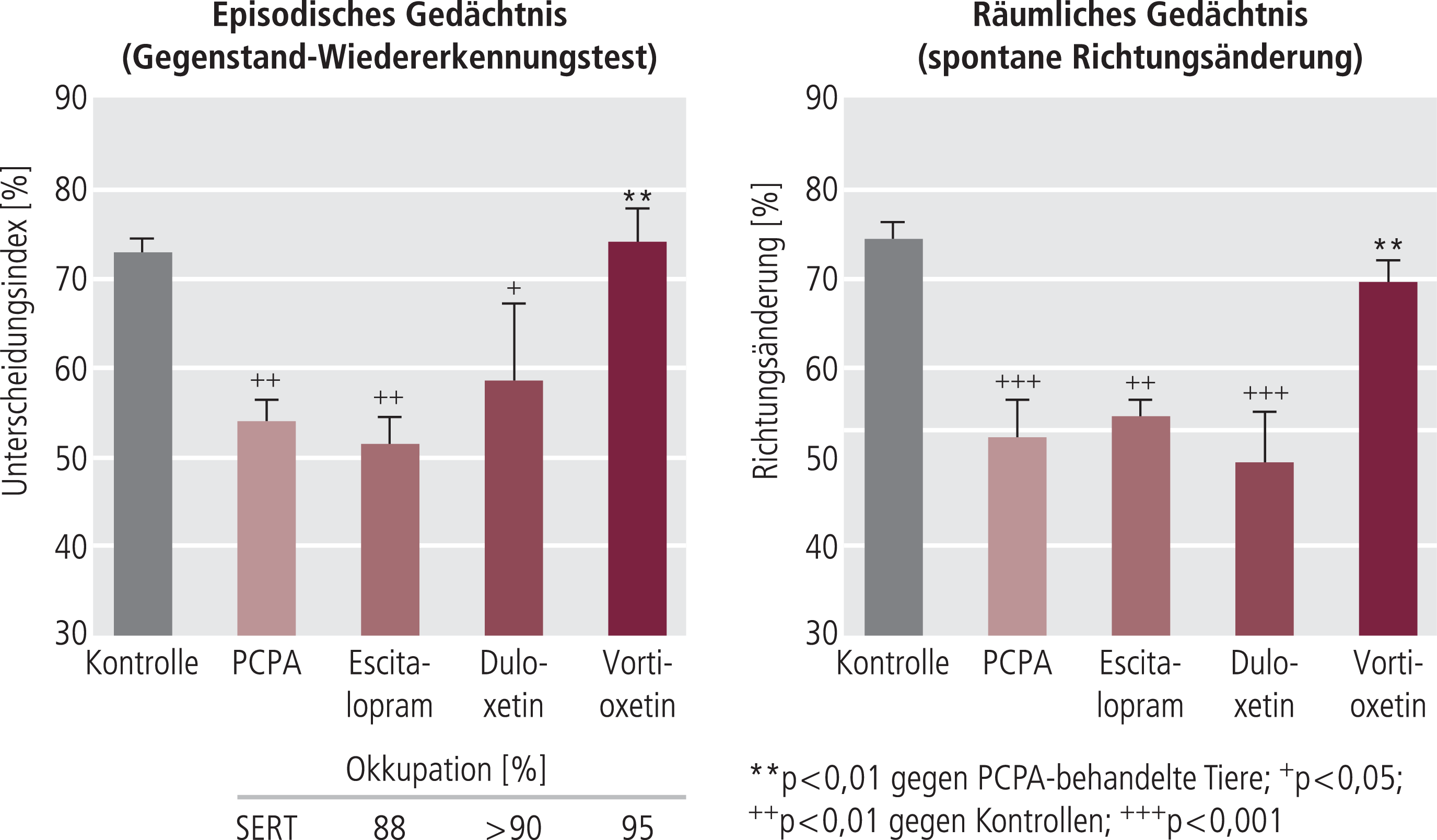
Abb. 9. Vortioxetin, nicht aber Escitalopram oder Duloxetin verbessern das Erinnerungsvermögen von Ratten nach Tryptophandepletion (TPD). Im Gegenstand-Wiedererkennungstest zeigte Vortioxetin einen sehr deutlichen Effekt, Escitalopram war unwirksam, während Duloxetin einen schwachen, nicht signifikanten Effekt zeigte. Die Daten stammen aus der Publikation von Jensen et al. [46].
Eine Tryptophandeplation führt auch in einem anderen auf natürliches Neugierverhalten zurückgehenden Kognitionstest (spontane Richtungsänderung) zu einer Verschlechterung des Gedächtnisses, die wiederum durch Vortioxetin, nicht aber durch Escitalopram oder Duloxetin reduziert werden konnte (Tab. 3) [46]. Alle drei Substanzen haben eine Hemmung von SERT und eine damit verbundene Erhöhung der extrazellulären Serotoninkonzentration gemeinsam, die bei Vortioxetin, wahrscheinlich durch die zusätzliche 5-HT3-Blockade, deutlich größer ist als selbst bei Escitalopram [65, 70]. Unter den oben genannten Dosen war die Okkupation von SERT im Rattenhirn nicht unterschiedlich [46], sodass der spezifische Effekt von Vortioxetin auf die Kognition auf andere, zusätzliche pharmakologische Mechanismen zurückgehen muss, die im Anschluss dargestellt werden. Vortioxetin konnte auch Defizite des räumlichen Gedächtnisses bei älteren Mäusen aufheben, der SSRI Fluoxetin war hingegen weniger wirksam [52]. Die tierexperimentellen Daten zur Kognitionsverbesserung sind in Tabelle 3 zusammenfasst. Vorläufige Daten zeigen auch eine Kognitionsverbesserung nach Defiziten induziert durch Scopolamin oder NMDA-Antagonisten [87].
Tab. 3. Tierexperimentelle Studien über Effekte von Vortioxetin auf Lernen und Gedächtnis
|
Publikation |
Test |
Tiere (Dosis) |
Ergebnis |
|
Mork et al. |
|
|
|
|
Du Jardin et al. [24] |
|
|
|
|
Jensen et al. [46] |
|
|
|
|
Wallace et al. [92] |
|
|
|
|
Li et al. [52] |
|
|
|
|
Sanchez et al. [86] |
|
|
|
Zum Wirkungsmechanismus von Vortioxetin
Vortioxetin ist eine Substanz, die den Serotonin-Wiederaufnahmetransporter hemmt, allerdings mit zusätzlichen agonistischen, partialagonistischen oder antagonistischen Effekten an verschiedenen Serotoninrezeptoren (Abb. 5) [5]. Im Gegensatz zu den Trizyklika, wo auch Rezeptoren außerhalb der primären Targetsysteme beeinflusst werden (z.B. Histamin oder Acetylcholin), bezieht sich das Rezeptorprofil von Vortioxetin nur auf die serotonerge Neurotransmission [37, 85]. Sein Wirkungsmechanismus wird nach neueren Klassifikationen auch als „multimodal“ bezeichnet [37]. Aufgrund dieses rein „serotonergen“ Profils ist es nicht verwunderlich, dass Vortioxetin in den meisten, allerdings nicht allen verhaltenspharmakologischen Modellen aktiv ist, die als indikativ für anxiolytische und besonders antidepressive Effekte gelten [8, 30, 65] und in klinischen Untersuchungen eine gute Wirksamkeit bei depressiven Patienten gezeigt hat [26, 60]. Gerade bei den klinischen Prüfungen hat es sich darüber hinaus gezeigt, dass Vortioxetin eine deutliche Wirksamkeit auf kognitive Symptome zeigt. Die Erklärung liegt in dem zusätzlichen spezifischen Rezeptorprofil dieser neuartigen Substanz (Abb. 5). Die Substanz bindet sehr stark an den 5-HT3-Rezeptor, etwas schwächer an SERT, gefolgt von 5-HT1A-, 5-HT1D-, 5-HT1B- und 5-HT7-Rezeptor. Wahrscheinlich bedingt durch die starke Blockade des 5-HT3-Rezeptors ist die Erhöhung von Serotonin bei gleicher SERT-Okkupation stärker als bei SSRI, was auch in Kombinationsversuchen (Citalopram und Ondansetron) gezeigt wurde [66]. Über diesen indirekten Effekt auf die extrazelluläre Serotoninkonzentration hinaus ist es möglich, dass bei Vortioxetin auch 5-HT3-Antagonismus [9], 5-HT1A-Agonismus, 5-HT1B-Partialantagonismus [32], 5-HT1D- und 5-HT7-Antagonismus [75] direkt zu den antidepressiven Eigenschaften beitragen, wobei allerdings der kombinierte Effekt von SERT- und 5-HT3-Blockade dominierend zu sein scheint [8].
Bei den überlegenen Effekten von Vortioxetin auf kognitive Störungen bei depressiven Patienten mit möglicher funktioneller serotonerger Narbe oder bei Versuchstieren nach Tryptophandepletion scheint eine alleinige Serotoninerhöhung nicht auszureichen, wie bereits ausgeführt. Hier scheinen der 5-HT3-Antagonismus und der 5-HT1A-Agonismus primär verantwortlich zu sein, da die Effekte vom 5-HT3-Antagonisten Ondansedron und vom 5-HT1A-Agonisten Flesinoxan nachgeahmt werden konnten [24]. Eine prokognitive Wirkung beider Mechanismen wird auch durch andere direkte Befunde bestätigt [4, 22, 48]. Auch für antagonistische Effekte an 5-HT1B- und 5-HT7-Rezeptoren gibt es solche Daten, allerdings keine im direkten Vergleich zu Vortioxetin [19, 22, 27, 58, 75]. Wahrscheinlich bedingt durch dieses spezifische Rezeptorprofil zeigt Vortioxetin auch in Mikrodialyseuntersuchungen Veränderungen der extrazellulären Konzentrationen von Noradrenalin, Dopamin, L-Glutamat und GABA, die so nicht für den SSRI Escitalopram gesehen wurden [72]. Während die zusätzliche Erhöhung von extrazellulärem Noradrenalin und Dopamin eine Brücke zu Duloxetin schlägt, bleibt eine mögliche Rolle der Erhöhung von L-Glutamat noch spekulativ [71].
Abschließende Bewertung
Für die besondere Wirkung von Vortioxetin auf kognitive Symptome depressiver Patienten bzw. bei Versuchstieren nach experimentell induzierter serotonergen Dysfunktion scheint im Vergleich zu SSRI das zusätzliche serotonerge Rezeptorprofil, besonders der 5-HT1A-Agonismus, der 5-HT3-Antagonismus und der 5-HT7-Antagonismus, wichtiger zu sein als die stärkere Erhöhung der extrazellulären Serotoninkonzentration im Vergleich mit typischen SSRI oder selbst Escitalopram, da die negativen Effekte der Serotoninerhöhung durch die zusätzlichen Effekte auf diese Serotoninrezeptoren abgeblockt werden.
Interessenkonflikterklärung
WEM hat Vortrags- und Beraterhonorare von Dr. Willmar Schwabe GmbH & Co KG (Karlsruhe) und Lundbeck (Hamburg) erhalten sowie Forschungsunterstützung von Dr. Willmar Schwabe GmbH & Co KG (Karlsruhe).
Literatur
1. Amado-Boccara I, Gougoulis N, Poirier Littré MF, Galinowski A, et al. Effects of antidepressants on cognitive functions: a review. Neurosci Biobehav Rev 1995;19:479–93.
2. Antunes M, Biala G. The novel object recognition memory: neurobiology, test procedure, and its modifications. Cogn Process 2012;13:93–110.
3. Ardis TC, Cahir M, Elliott JJ, Bell R, et al. Effect of acute tryptophan depletion on noradrenaline and dopamine in the rat brain. J Psychopharmacol 2009;23:51–5.
4. Arnsten AF, Lin CH, Van Dyck CH, Stanhope KJ. The effects of 5-HT3 receptor antagonists on cognitive performance in aged monkeys. Neurobiol Aging 1997;18:21–8.
5. Bang-Andersen B, Ruhland T, Jørgensen M, Smith G, et al. Discovery of 1-[2-(2,4-dimethylphenylsulfanyl)phenyl]piperazine (Lu AA21004): a novel multimodal compound for the treatment of major depressive disorder. J Med Chem 2011;54:3206–21.
6. Bell C, Abrams J, Nutt D. Tryptophan depletion and its implications for psychiatry. Br J Psychiatry 2001;178:399–405.
7. Berigan TR. Atomoxetine used adjunctively with selective serotonin reuptake inhibitors to treat depression. Prim Care Companion J Clin Psychiatry 2004;6:93–4.
8. Bétry C, Pehrson AL, Etiévant A, Ebert B, et al. The rapid recovery of 5-HT cell firing induced by the antidepressant vortioxetine involves 5-HT3 receptor antagonism. Int J Neuropsychopharmacol 2013;16:1115–27.
9. Bétry C, Etiévant A, Oosterhof C, Ebert B, et al. Role of 5-HT3 receptors in the antidepressant response. Pharmaceuticals 2011;4:603–29.
10. Blier P, El Mansari M. Serotonin and beyond: therapeutics for major depression. Philos Trans R Soc Lond B Biol Sci 2013;368:20120536.
11. Blier P. Neurotransmitter targeting in the treatment of depression. J Clin Psychiatry 2013;74(Suppl 2):19–24.
12. Blier P. Rational site-directed pharmacotherapy for major depressive disorder. Int J Neuropsychopharmacol 2014;17:997–1008.
13. Bora E, Harrison BJ, Yücel M, Pantelis C. Cognitive impairment in euthymic major depressive disorder: a meta-analysis. Psychol Med 2013;43:2017–26.
14. Borkowska A, Drozdz W, Ziółkowska-Kochan M, Rybakowski J. Enhancing effect of mirtazapine on cognitive functions associated with prefrontal cortex in patients with recurrent depression. Neuropsychopharmacol Hung 2007;9:131–6.
15. Bymaster FP, Katner JS, Nelson DL, Hemrick-Luecke SK, et al. Atomoxetine increases extracellular levels of norepinephrine and dopamine in prefrontal cortex of rat: a potential mechanism for efficacy in attention deficit/hyperactivity disorder. Neuropsychopharmacology 2002;27:699–711.
16. Carlini VP, Poretti MB, Rask-Andersen M, Chavan RA, et al. Differential effects of fluoxetine and venlafaxine on memory recognition: possible mechanisms of action. Prog Neuropsychopharmacol Biol Psychiatry 2012;38:159–67.
17. Castrén E, Hen R. Neuronal plasticity and antidepressant actions. Trends Neurosci 2013;36:259–67.
18. Chenu F, El Mansari M, Blier P. Electrophysiological effects of repeated administration of agomelatine on the dopamine, norepinephrine, and serotonin systems in the rat brain. Neuropsychopharmacology 2013;38:275–84.
19. Cifariello A, Pompili A, Gasbarri A. 5-HT7 receptors in the modulation of cognitive processes. Behav Brain Res 2008;195:171–9.
20. Clark L, Chamberlain SR, Sahakian BJ. Neurocognitive mechanisms in depression: implications for treatment. Annu Rev Neurosci 2009;32:57–74.
21. Connolly KR, Thase ME. Emerging drugs for major depressive disorder. Expert Opin Emerg Drugs 2012;17:105–26.
22. Cowen P, Sherwood AC. The role of serotonin in cognitive function: evidence from recent studies and implications for understanding depression. J Psychopharmacol 2013;27:575–83.
23. Cowen PJ. Serotonin and depression: pathophysiological mechanism or marketing myth? Trends Pharmacol Sci 2008;29:433–6.
24. Du Jardin KG, Jensen JB, Sanchez C, Pehrson AL. Vortioxetine dose-dependently reverses 5-HT depletion-induced deficits in spatial working and object recognition memory: a potential role for 5-HT1A receptor agonism and 5-HT3 receptor antagonism. Eur Neuropsychopharmacol 2014;24:160–71.
25. Ferguson JM, Wesnes KA, Schwartz GE. Reboxetine versus paroxetine versus placebo: effects on cognitive functioning in depressed patients. Int Clin Psychopharmacol 2003;18:9–14.
26. Garnock-Jones KP. Vortioxetine: a review of its use in major depressive disorder. CNS Drugs 2014;28:855–74.
27. Gasbarri A, Pompili A. Serotonergic 5-HT7 receptors and cognition. Rev Neurosci 2014;25:311–23.
28. Greer TL, Sunderajan P, Grannemann BD, Kurian BT, et al. Does duloxetine improve cognitive function independently of its antidepressant effect in patients with major depressive disorder and subjective reports of cognitive dysfunction? Depress Res Treat 2014;2014: 627863.
29. Gualtieri CT, Johnson LG, Benedict KB. Neurocognition in depression: patients on and off medication versus healthy comparison subjects. J Neuropsychiatry Clin Neurosci 2006;18:217–25.
30. Guilloux JP, Mendez-David I, Pehrson A, Guiard BP, et al. Antidepressant and anxiolytic potential of the multimodal antidepressant vortioxetine (Lu AA21004) assessed by behavioural and neurogenesis outcomes in mice. Neuropharmacology 2013;73:147–59.
31. Haddad AD, Williams JM, McTavish SF, Harmer CJ. Low-dose tryptophan depletion in recovered depressed women induces impairments in autobiographical memory specificity. Psychopharmacology (Berl) 2009;207:499–508.
32. Hamon M, Blier P. Monoamine neurocircuitry in depression and strategies for new treatments. Prog Neuropsychopharmacol Biol Psychiatry 2013;45:54–63.
33. Hasselbalch BJ, Knorr U, Kessing LV. Cognitive impairment in the remitted state of unipolar depressive disorder: a systematic review. J Affect Disord 2011;134:20–31.
34. Hasselbalch BJ, Knorr U, Hasselbalch SG, Gade A, et al. Cognitive deficits in the remitted state of unipolar depressive disorder. Neuropsychology 2012;26:642–51.
35. Hasselbalch BJ, Knorr U, Hasselbalch SG, Gade A, et al. The cumulative load of depressive illness is associated with cognitive function in the remitted state of unipolar depressive disorder. Eur Psychiatry 2013;28:349–55.
36. Hayward G, Goodwin GM, Cowen PJ, Harmer CJ. Low-dose tryptophan depletion in recovered depressed patients induces changes in cognitive processing without depressive symptoms. Biol Psychiatry 2005;57:517–24.
37. Hellweg R, Goemann C. Effekt von Vortioxetin auf depressive Symptome einer Major Depression. Psychopharmakotherapie 2014;21:134–41.
38. Herrera-Guzmán I, Gudayol-Ferré E, Herrera-Abarca JE, Herrera-Guzmán D, et al. Major depressive disorder in recovery and neuropsychological functioning: effects of selective serotonin reuptake inhibitor and dual inhibitor depression treatments on residual cognitive deficits in patients with major depressive disorder in recovery. J Affect Disord 2010;123:341–50.
39. Herrera-Guzmán I, Gudayol-Ferré E, Herrera-Guzmán D, Guàrdia-Olmos J, et al. Effects of selective serotonin reuptake and dual serotonergic-noradrenergic reuptake treatments on memory and mental processing speed in patients with major depressive disorder. J Psychiatr Res 2009;43:855–63.
40. Herrera-Guzmán I, Gudayol-Ferré E, Lira-Mandujano J, Herrera-Abarca J, et al. Cognitive predictors of treatment response to bupropion and cognitive effects of bupropion in patients with major depressive disorder. Psychiatry Res 2008;160:72–82.
41. Herrera-Guzmán I, Herrera-Abarca JE, Gudayol-Ferré E, Herrera-Guzmán D, et al. Effects of selective serotonin reuptake and dual serotonergic-noradrenergic reuptake treatments on attention and executive functions in patients with major depressive disorder. Psychiatry Res 2010;177:323–9.
42. Izquierdo A, Carlos K, Ostrander S, Rodriguez D, et al. Impaired reward learning and intact motivation after serotonin depletion in rats. Behav Brain Res 2012;233:494–9.
43. Jaeger J, Berns S, Uzelac S, Davis-Conway S. Neurocognitive deficits and disability in major depressive disorder. Psychiatry Res 2006;145:39–48.
44. Jans LA, Korte-Bouws GA, Korte SM, Blokland A. The effects of acute tryptophan depletion on affective behaviour and cognition in Brown Norway and Sprague Dawley rats. J Psychopharmacol 2010;24:605–14.
45. Jenkins TA, Elliott JJ, Ardis TC, Cahir M, et al. Tryptophan depletion impairs object-recognition memory in the rat: reversal by risperidone. Behav Brain Res 2010;208:479–83.
46. Jensen JB, du Jardin KG, Song D, Budac D, et al. Vortioxetine, but not escitalopram or duloxetine, reverses memory impairment induced by central 5-HT depletion in rats: evidence for direct 5-HT receptor modulation. Eur Neuropsychopharmacol 2014;24:148–59.
47. Katona C, Hansen T, Olsen CK. A randomized, double-blind, placebo-controlled, duloxetine-referenced, fixed-dose study comparing the efficacy and safety of Lu AA21004 in elderly patients with major depressive disorder. Int Clin Psychopharmacol 2012;27:215–23.
48. King MV, Marsden CA, Fone KC. A role for the 5-HT1A, 5-HT4 and 5-HT6 receptors in learning and memory. Trends Pharmacol Sci 2008;29:482–92.
49. Lapiz-Bluhm MD, Soto-Piña AE, Hensler JG, Morilak DA. Chronic intermittent cold stress and serotonin depletion induce deficits of reversal learning in an attentional set-shifting test in rats. Psychopharmacology (Berl) 2009;202:329–41.
50. Laux G, Goemann C. „Unmet needs“ in der Antidepressiva-Therapie. Psychopharmakotherapie 2014;21:7–11.
51. Leiser SC, Pehrson AL, Robichaud PJ, Sanchez C. Multimodal antidepressant vortioxetine increases frontal cortical oscillations unlike escitalopram and duloxetine – a quantitative EEG study in rats. Br J Pharmacol 2014;171:4255–72.
52. Li Y, Sanchez C, Gulinello M. Memory impairment in old mice is differentially sensitive to different classes of antidepressants. Eur Neuropsychopharmacol 2013;23(Suppl 2):S282.
53. Mahableshwarkar A, et al. Efficacy of vortioxetine on cognitive function in adult patients with major depressive disorder: Results of a randomized, double-blind, active-referenced, placebo-controlled trial. Abstract 016, presented at CINP; LP-02-Late Breaking Poster Session.
54. McClintock SM, Husain MM, Wisniewski SR, Nierenberg AA, et al. Residual symptoms in depressed outpatients who respond by 50% but do not remit to antidepressant medication. J Clin Psychopharmacol 2011;31:180–6. Erratum in: J Clin Psychopharmacol 2013;33:765.
55. McIntyre RS, Cha DS, Soczynska JK, Woldeyohannes HO, et al. Cognitive deficits and functional outcomes in major depressive disorder: determinants, substrates, and treatment interventions. Depress Anxiety 2013;30:515–27.
56. McIntyre RS, Lophaven S, Olsen CK. A randomized, double-blind, placebo-controlled study of vortioxetine on cognitive function in depressed adults. Int J Neuropsychopharmacol 2014;17:1557–67.
57. Mendelsohn D, Riedel WJ, Sambeth A. Effects of acute tryptophan depletion on memory, attention and executive functions: a systematic review. Neurosci Biobehav Rev 2009;33:926–52.
58. Meneses A. Memory formation and memory alterations: 5-HT6 and 5-HT7 receptors, novel alternative. Rev Neurosci 2014;25:325–56.
59. Merens W, Booij L, Haffmans PJ, van der Does A. The effects of experimentally lowered serotonin function on emotional information processing and memory in remitted depressed patients. J Psychopharmacol 2008;22:653–62.
60. Messer T, Goemann C. Vortioxetin – ein multimodales Antidepressivum. Psyschopharmakotherapie 2014;21:142–9.
61. Millan MJ, Agid Y, Brüne M, Bullmore ET, et al. Cognitive dysfunction in psychiatric disorders: characteristics, causes and the quest for improved therapy. Nat Rev Drug Discov 2012;11:141–68.
62. Möller H-J. Realität der ambulanten psychotherapeutischen Versorgung im Kontrast zur Nationalen Versorgungsleitlinie Depression. Psyschopharmakotherapie 2014;21:12–20.
63. Monleón S, Vinader-Caerols C, Arenas MC, Parra A. Antidepressant drugs and memory: insights from animal studies. Eur Neuropsychopharmacol 2008;18:235–48.
64. Morimoto SS, Alexopoulos GS. Cognitive deficits in geriatric depression: clinical correlates and implications for current and future treatment. Psychiatr Clin North Am 2013;36:517–31.
65. Mørk A, Pehrson A, Brennum LT, Nielsen SM, et al. Pharmacological effects of Lu AA21004: a novel multimodal compound for the treatment of major depressive disorder. J Pharmacol Exp Ther 2012;340:666–75.
66. Mørk A, Montezinho LP, Miller S, Trippodi-Murphy C, et al. Vortioxetine (Lu AA21004), a novel multimodal antidepressant, enhances memory in rats. Pharmacol Biochem Behav 2013;105:41–50.
67. Müller WE, Eckert A. Psychopharmakotherapie – Pharmakologische Grundlagen (2011). In: Möller HJ, Laux G, Kapfhammer KP (Hrsg.). Psychiatrie, Psychosomatik, Psychotherapie. Band 1. Berlin, Heidelberg: Springer, 2010: 683–730.
68. Olivier JD, Jans LA, Korte-Bouws GA, Korte SM, et al. Acute tryptophan depletion dose dependently impairs object memory in serotonin transporter knockout rats. Psychopharmacology (Berl) 2008;200:243–54.
69. Otte C. Depression und kognitive Dysfunktion. Psychopharmakotherapie 2014;21:40–9.
70. Pehrson AL, Cremers T, Bétry C, van der Hart MG, et al. Lu AA21004, a novel multimodal antidepressant, produces regionally selective increases of multiple neurotransmitters – a rat microdialysis and electrophysiology study. Eur Neuropsychopharmacol 2013;23:133–45.
71. Pehrson AL, Sanchez C. Serotonergic modulation of glutamate neurotransmission as a strategy for treating depression and cognitive dysfunction. CNS Spectr 2014;19:121–33.
72. Pehrson AL, Leiser SC, Gulinello M, Dale E, et al. Treatment of cognitive dysfunction in major depressive disorder – a review of the preclinical evidence for efficacy of selective serotonin reuptake inhibitors, serotonin-norepinephrine reuptake inhibitors and the multimodal-acting antidepressant vortioxetine. Eur J Pharmacol 2015;753:19–31.
73. Pittenger C, Duman RS. Stress, depression, and neuroplasticity: a convergence of mechanisms. Neuropsychopharmacology 2008;33:88–109.
74. Porter RJ, Phipps AJ, Gallagher P, Scott A, et al. Effects of acute tryptophan depletion on mood and cognitive functioning in older recovered depressed subjects. Am J Geriatr Psychiatry 2005;13:607–15.
75. Pouzet B. SB-258741: a 5-HT7 receptor antagonist of potential clinical interest. CNS Drug Rev 2002;8:90–100.
76. Raskin J, Wiltse CG, Siegal A, Sheikh J, et al. Efficacy of duloxetine on cognition, depression, and pain in elderly patients with major depressive disorder: an 8-week, double-blind, placebo-controlled trial. Am J Psychiatry 2007;164:900–9.
77. Reppermund S, Zihl J, Lucae S, Horstmann S, et al. Persistent cognitive impairment in depression: the role of psychopathology and altered hypothalamic-pituitary-adrenocortical (HPA) system regulation. Biol Psychiatry 2007;62:400–6.
78. Reppermund S, Ising M, Lucae S, Zihl J. Cognitive impairment in unipolar depression is persistent and non-specific: further evidence for the final common pathway disorder hypothesis. Psychol Med 2009;39:603–14.
79. Riedel WJ, Sobczak S, Schmitt JA. Tryptophan modulation and cognition. Adv Exp Med Biol 2003;527:207–13. Review.
80. Riedel WJ. Cognitive changes after acute tryptophan depletion: what can they tell us? Psychol Med 2004;34:3–8.
81. Robbins TW, Arnsten AF. The neuropsychopharmacology of fronto-executive function: monoaminergic modulation. Annu Rev Neurosci 2009;32:267–87.
82. Rock PL, Roiser JP, Riedel WJ, Blackwell AD. Cognitive impairment in depression: a systematic review and meta-analysis. Psychol Med 2013 Oct 29:1–12.
83. Roiser JP, Müller U, Clark L, Sahakian BJ, et al. The effects of acute tryptophan depletion and serotonin transporter polymorphism on emotional processing in memory and attention. Int J Neuropsychopharmacol 2007;10:449–61.
84. Roiser JP, Elliott R, Sahakian BJ. Cognitive mechanisms of treatment in depression. Neuropsychopharmacology 2012;37:117–36.
85. Sanchez C, Asin KE, Artigas F. Vortioxetine, a novel antidepressant with multimodal activity: Review of preclinical and clinical data. Pharmacol Ther 2014 Jul 9. pii: S0163–7258(14)00127–2.
86. Sanchez C, Pehrson AL, Li Y, Haddjeri N, et al. The multimodal antidepressant vortioxetine restores cognitive function in preclinical models across several cognitive domains. Poster, 53rd ACNP Meeting, December 2014, Phoenix (USA).
87. Snyder HR. Major depressive disorder is associated with broad impairments on neuropsychological measures of executive function: a meta-analysis and review. Psychol Bull 2013;139:81–132.
88. Swanson CJ, Perry KW, Koch-Krueger S, Katner J, et al. Effect of the attention deficit/hyperactivity disorder drug atomoxetine on extracellular concentrations of norepinephrine and dopamine in several brain regions of the rat. Neuropharmacology 2006;50:755–60.
89. Trivedi MH, Greer TL. Cognitive dysfunction in unipolar depression: implications for treatment. J Affect Disord 2014;152–154: 19–27.
90. Uher R, Perlis RH, Henigsberg N, Zobel A, et al. Depression symptom dimensions as predictors of antidepressant treatment outcome: replicable evidence for interest-activity symptoms. Psychol Med 2012;42:967–80.
91. Wagner S, Doering B, Helmreich I, Lieb K, et al. A meta-analysis of executive dysfunctions in unipolar major depressive disorder without psychotic symptoms and their changes during antidepressant treatment. Acta Psychiatr Scand 2012;125:281–92.
92. Wallace A, Pehrson AL, Sánchez C, Morilak DA. Vortioxetine restores reversal learning impaired by 5-HT depletion or chronic intermittent cold stress in rats. Int J Neuropsychopharmacol 2014;17:1695–706.
93. Willner P, Scheel-Krüger J, Belzung C. The neurobiology of depression and antidepressant action. Neurosci Biobehav Rev 2013;37:2331–71.
94. Zajecka J, Kornstein SG, Blier P. Residual symptoms in major depressive disorder: prevalence, effects, and management. J Clin Psychiatry 2013;74:407–14.
Univ.-Prof. Dr. Walter E. Müller, Höhenstraße 49A, 67550 Worms
Antidepressant drugs and cognitive dysfunction: the role of vortioxetine
Cognitive dysfunctions are frequent symptoms of depression which only partly respond to antidepressant treatment but persist frequently over remission of other symptoms. The assumption that the mechanisms underlying cognitive dysfunction or other symptoms of depression might differ was confirmed by studies using tryptophan-depletion where cognition was much more sensitive than mood. Although this might indicate an association between low serotonin and cognitive dysfunction in depression, SSRI’s, which elevate serotonin, are not very active in reducing cognitive dysfunction in depression or in respective animal models. However, such effects are clearly present for the new multimodal antidepressant vortioxetine which not only elevates serotonin but also shows substantial affinity for many serotonin receptors. Some of these receptors, mainly 5-HT3 and 5-HT7 have negative effects on cognition once activated by serotonin. These negative effects of serotonin elevation e.g. in case of vortioxetine are blocked by its own receptor profile.
Key words: Antidepressant drugs, cognitive improvement, vortioxetine, animal data
Psychopharmakotherapie 2015; 22(04)